Limfosit B:
Limfosit B mendapatkan namanya dari tempat pematangannya, bursa Fabricious, pada burung.
Limfosit B diproduksi dari sel induk hematopoietik di sumsum tulang orang dewasa. Limfosit B matang yang dilepaskan ke dalam sirkulasi dari sumsum tulang berada dalam keadaan istirahat atau perawan. Limfosit B yang beristirahat tidak mengeluarkan antibodi. Alih-alih, sel B yang beristirahat mengekspresikan imunoglobulin permukaan (sig) pada membrannya.
Perkembangan sel B:
Diperkirakan 5 x sel B/hari diproduksi di sumsum tulang. Namun, hanya 10 persen dari sel B ini yang dilepaskan ke dalam sirkulasi. Sisa 90 persen sel B yang diproduksi setiap hari mati di dalam sumsum tulang. Selama perkembangannya di sumsum tulang, sejumlah besar sel B yang sedang berkembang dibunuh oleh proses yang dikenal sebagai seleksi negatif sel B. Sel B yang berkembang mampu bereaksi dengan self-antigen dibunuh di sumsum tulang.
Respon Kekebalan Humoral:
Ada dua lengan kekebalan yang didapat yang disebut kekebalan humoral dan kekebalan yang dimediasi sel. Sel B terlibat dalam imunitas humoral. Setelah pengikatan antigen, sel B diaktifkan. Sel B yang teraktivasi membelah berulang kali untuk menghasilkan sel plasma dan sel B memori. Sel plasma mengeluarkan antibodi dan antibodi adalah molekul penting dari respon imun humoral.
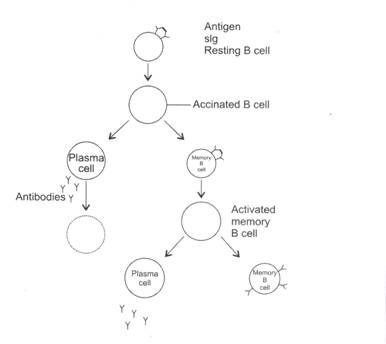
Gambar 8.1: Aktivasi sel B.
Sel B yang sedang beristirahat diaktifkan oleh pengikatan imunoglobulin permukaannya (slg) dengan antigen. Sel B yang teraktivasi membelah berulang kali. Beberapa sel anak menjadi sel efektor (plasma) dan yang lainnya menjadi sel B memori. Sel plasma mengeluarkan antibodi.
Sel plasma memiliki umur yang pendek dan mati dalam beberapa hari. Sel B memori dalam keadaan istirahat dan mereka tetap berada di inang selama berbulan-bulan hingga bertahun-tahun. Ketika sel B memori kebetulan menghubungi antigen serupa (yang menginduksi produksi sel B memori dari sel B yang diaktifkan), sel B memori akan diaktifkan. Sel B yang teraktivasi membelah berkali-kali untuk menghasilkan sel efektor (plasma) dan sel B memori
B Aktivasi Limfosit:
Limfosit B dewasa yang dilepaskan dari sumsum tulang dalam keadaan istirahat dan tidak mengeluarkan antibodi. Sel B istirahat, yang tidak menghubungi antigen mati dalam beberapa hari. Sedangkan, sel B istirahat, yang berikatan dengan antigen melalui imunoglobulin permukaan sel B (sigs) menjadi aktif. Sel B yang teraktivasi membelah dan menghasilkan sel plasma dan sel B memori.
Aktivasi sel B membutuhkan dua sinyal dan sitokin dari sel TH :
saya. Sinyal pertama disediakan oleh pengikatan antigen ke permukaan imunoglobulin [sIgs; juga disebut membrane immunoglobulin’s (migs)] pada permukaan sel B.
- Kontak sel-ke-sel antara sel B dan sel TH memberikan sinyal kedua yang diperlukan untuk aktivasi sel B.
Pengikatan antigen ke imunoglobulin permukaan memberikan sinyal pertama dan memulai aktivasi sel B. Antigen berikatan dengan sIg pada sel B dan menyilangkan sIg.
sebuah†”
Kompleks sIg-antigen diinternalisasi ke dalam sel B melalui endositosis (disebut sebagai endositosis yang dimediasi reseptor atau endositosis yang dimediasi imunoglobulin).
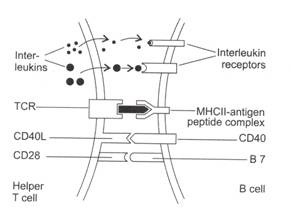
Gambar 8.2: Aktivasi sel T Helper dan sel B.
Tanda pada sel B yang beristirahat berikatan dengan antigen dan antigen diinternalisasi. Enzim lisosom memecah antigen menjadi peptida kecil dan membentuk kompleks menjadi antigen MHC kelas II. Kompleks peptida ll-antlgen kelas MHC diekspresikan pada permukaan sel B.
Pengikatan berikut terjadi antara molekul pada sel B dan sel T:
- Reseptor sel T (TCR) sel T helper berikatan dengan kompleks peptida antigen MHC kelas II pada sel B.
- Pengikatan antara molekul CD40L (pada sel T) dengan CD40 (pada sel B) memberikan sinyal kedua untuk aktivasi sel B.
- Molekul CD28 (pada sel T) berinteraksi dengan B7 (pada sel B) dan memberikan sinyal rangsangan yang diperlukan ke sel B. Selanjutnya, sitokin yang disekresikan oleh sel T berikatan dengan reseptor sitokin sel B pada sel B dan membantu aktivasi sel B.
Antigen yang diinternalisasi diproses menjadi peptida antigen melalui jalur endositik. Enzim lisosom sel B membelah antigen menjadi peptida antigen pendek.
sebuah†”
Peptida antigen pendek dikomplekskan ke molekul MHC kelas II. Kompleks peptida Il-antigen kelas MHC diangkut ke permukaan sel B dan diekspresikan pada permukaan sel B.
sebuah†”
Kompleks peptida antigen MHC kelas II pada sel B dipresentasikan ke sel TH . Biasanya diperlukan waktu sekitar 30-60 menit bagi sel B untuk memproses dan menyajikan antigen ke sel TH . (Pengikatan antigen ke sIgs pada sel B juga menginduksi sel B untuk mengekspresikan sejumlah besar molekul kelas II dan molekul B7 pada permukaan sel B. Induksi ekspresi molekul MHC kelas II membantu dalam presentasi yang lebih baik dari kompleks peptida Il-antigen kelas MHC ke sel TH .)
sebuah†”
Reseptor sel T (TCR) sel T H berikatan dengan kompleks MHC kelas Il-antigen pada sel B dan membentuk konjugat TB.
sebuah†”
Sinyal kedua untuk aktivasi sel B disediakan oleh kontak langsung antara beberapa molekul pada sel TH dan sel B. Interaksi antara molekul CD40 pada sel B dan CD40L (ligan CD40) pada sel TH memberikan sinyal kedua yang diperlukan untuk aktivasi sel B. Secara umum, pengikatan antigen ke imunoglobulin permukaan dan stimulasi CD40L bekerja secara sinergis untuk memicu aktivasi sel B.
sebuah†”
Juga, molekul B7 (pada sel B) berinteraksi dengan molekul CD28 (pada sel TH ). Interaksi sel-ke-sel ini memberikan sinyal stimulasi yang dibutuhkan oleh sel TH untuk aktivasi sel T. Sel TH yang teraktivasi , pada gilirannya mengeluarkan banyak sitokin seperti IL-2, IL-4, IL-5, dan lain-lain.
sebuah†”
Sel B yang teraktivasi mengekspresikan reseptor permukaan untuk berbagai sitokin, seperti IL-2, IL-4, IL-5, dan lain-lain. Sitokin yang disekresikan oleh sel T H berikatan dengan reseptornya masing-masing pada sel B dan membantu proliferasi dan diferensiasi sel B lebih lanjut.
sebuah†”
Sel B yang diaktifkan membelah berkali-kali. Beberapa sel anak menjadi sel plasma dan yang lainnya menjadi sel B memori.
Selain sIgs, beberapa molekul lain pada permukaan sel B juga berperan penting dalam aktivasi sel B. Mereka adalah rantai Ig-a/Ig-P, kompleks koreseptor sel B, dan molekul CD22 pada permukaan sel B.
Reseptor Sel B dan Aktivasi Sel B:
Setiap sig pada sel B dikaitkan dengan dua polipeptida heterodimer Ig-α/Igβ yang mentransduksi sinyal untuk membentuk reseptor sel B (BCR). Rantai polipeptida Ig-α dan Ig-β memiliki ekor sitoplasma yang panjang. Ekor sitoplasma dari kedua rantai Ig-α dan Ig-β mengandung motif 18-residu yang disebut motif aktivasi berbasis imuno-reseptor tirosin (ITAM).
Pengikatan antigen dan ikatan silang dari sigs memberikan stimulus awal untuk aktivasi sel B. Stimulus yang dihasilkan oleh ikatan silang sIgs ditransduksi ke dalam sel B oleh ekor sitoplasma Ig-α/ Ig-β. Sinyal aktivasi sel B dimediasi oleh protein tirosin kinase (PTK). Proses transduksi sinyal mengarah pada pembentukan faktor transkripsi aktif. Faktor transkripsi merangsang transkripsi gen spesifik dalam inti sel B.
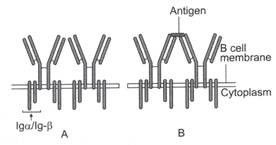
Gambar 8.3: Reseptor sel B.
(A) Imunoglobulin permukaan (sIg) dan dua rantai polipeptida Ig-α/lg-β pentransduksi sinyal membentuk reseptor sel B pada membran sel B. Rantai Ig-α/lg-β memiliki ekor sitoplasma yang panjang. Ekor sitoplasma mengandung motif 18-residu yang disebut motif aktivasi berbasis imuno-reseptor tirosin (ITAM). (B) Inisiasi aktivasi sel B. Antigen mengikat dan menyilangkan sIg yang berdekatan pada sel B. Pengikatan antigen dengan sIgs memberikan sinyal awal untuk aktivasi sel B. Setelah menghubungkan tanda silang, ITAM berinteraksi dengan banyak anggota keluarga Src dari tirosin kinase (Fyn, BIk, dan Lck) dan mengaktifkan kinase
Kompleks Koreseptor sel B dan Aktivasi sel B:
Kompleks koreseptor sel B terdiri dari tiga rantai protein yang disebut CD19, CR2 (CD21), dan CD81 (TAPA-1) (Gbr. 8.4). CD19 memiliki ekor sitoplasma yang panjang dan tiga domain lipatan Ig ekstra seluler. CR2 (reseptor komplemen 2) bertindak sebagai reseptor untuk C3d, produk penguraian yang terbentuk selama aktivasi komplemen. CD81 adalah rantai polipeptida sparming membran.
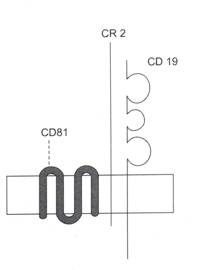
Gambar 8.4: koreseptor sel B.
Tiga rantai polipeptida membran sel B bersama-sama membentuk koreseptor sel B. GDI 9 memiliki ekor sitoplasma yang panjang dan tiga domain lipatan Ig ekstraseluler. CR2 (reseptor pelengkap) memiliki ekor sitoplasma yang pendek. Bagian ekstraseluler CR2 bertindak sebagai reseptor untuk fragmen komplemen C3d. CD81 adalah membran yang mencakup rantai polipeptida
Antibodi yang terbentuk melawan antigen berikatan dengan antigen spesifik.
sebuah†”
Pengikatan antibodi dengan antigen mengaktifkan jalur komplemen klasik dan menyebabkan pengendapan C3d pada antigen.
sebuah†”
Ketika antigen dalam kompleks antigen-antibodi berikatan dengan sIgs sel B, molekul CR2 yang berdekatan (pada sel B) berikatan dengan C3d pada antigen (CR2 bertindak sebagai reseptor untuk C3d). Jadi, sIgs dan koreseptor sel B dijembatani satu sama lain melalui kompleks antigen-antibodi (Gambar 8.5).
sebuah†”
Ikatan silang sIgs dengan ko-reseptor memungkinkan rantai CD19 (dari ko-reseptor sel B) untuk berinteraksi dengan rantai Ig-a / Ig-β rantai reseptor sel B. Kompleks ko-reseptor berfungsi untuk memperkuat sinyal pengaktifan yang ditransmisikan melalui BCR.
CD22 dan Sinyal Negatif untuk Aktivasi sel B:
Sel B juga mengekspresikan molekul yang disebut CD22 di permukaannya. CD22 secara konstitutif terkait dengan reseptor sel B dalam istirahat sel B. CD22 mengirimkan sinyal negatif yang membuat sel B lebih sulit untuk diaktifkan.
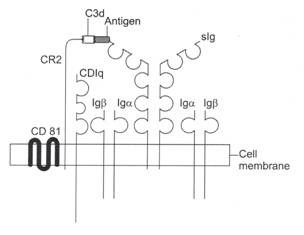
Gambar 8.5: Reseptor dan koreseptor sel B selama aktivasi sel B .
Antigen mengikat dan menyilangkan sigs. ITAM dari Ig-α/lg-β berinteraksi dengan anggota keluarga Src dari tirosin kinase dan mengaktifkan kinase. Rantai CR2 kompleks koreseptor sel B bertindak sebagai reseptor untuk C3d dan berikatan dengan C3d pada permukaan antigen. Pengikatan CR2 dengan C3d menyebabkan fosforilasi CD19. Keluarga Src dari tirosin kinase Lyn berikatan dengan CD19 terfosforilasi. Kompleks ko-reseptor memperkuat sinyal pengaktifan yang ditransmisikan melalui reseptor sel B
Sel Plasma dan Antibodi:
Ketika sel B diaktifkan, sel B yang diaktifkan membelah berulang kali. Beberapa sel yang terbagi menjadi sel plasma sementara yang lain menjadi sel B memori. Sel plasma mengeluarkan antibodi. Sel plasma berbentuk bulat atau elips. Sitoplasma berlimpah dan mungkin memiliki karakter granular. Nukleus kecil dalam kaitannya dengan ukuran sel. Nukleus ditempatkan secara eksentrik dan mengandung massa kromatin yang padat, seringkali tersusun dengan gaya roda. Sel plasma tidak bereplikasi.
Mereka hidup hanya beberapa hari dan kemudian mereka mati dengan proses yang disebut kematian sel terprogram. Sel plasma dapat mengeluarkan ribuan molekul antibodi per menit. Antibodi awal yang disekresikan oleh sel plasma sebagai respons terhadap antigen selalu milik kelas IgM. Biasanya respons humoral berkurang setelah berkurangnya tantangan antigenik karena sel plasma penghasil antibodi tidak berumur panjang.
Antibodi yang disekresikan oleh sel plasma adalah molekul polipeptida empat rantai berbentuk ‘Y’. Wilayah Fab antibodi berikatan dengan antigen spesifiknya. Antibodi dari sel plasma hanya mengikat antigen yang bertanggung jawab untuk produksinya (dengan mengaktifkan sel B dan akibatnya, mengarah pada perkembangan sel plasma). Antibodi spesifik untuk antigen tertentu, karena antibodi tidak berikatan dengan antigen lain.
Antibodi juga dikatakan sebagai molekul dwi-fungsional. Fungsi utama antibodi adalah mengikat antigen spesifiknya melalui daerah Fab-nya. Pengikatan antibodi terhadap antigen mengarah pada berfungsinya wilayah Fc antibodi (dan fungsi yang dimediasi oleh wilayah Fc dikatakan sebagai fungsi sekunder).
Memori B Limfosit:
Setelah aktivasi sel B, sel B yang diaktifkan membelah untuk menghasilkan dua kelompok sel yang disebut sel plasma dan sel B memori. Sel B memori tidak segera mengeluarkan antibodi. Mereka tetap dalam keadaan istirahat selama berbulan-bulan hingga bertahun-tahun. Faktanya, pada orang dewasa, kelenjar getah bening dikemas dengan sel B memori. Sel B memori di kelenjar getah bening sedang menunggu kontak dengan antigen spesifiknya.
Setelah kontaknya dengan antigen spesifik, sel B memori menjadi aktif. Sel B memori yang teraktivasi membelah dan menghasilkan sel plasma dan sel B memori (Gambar 8.1). Sel plasma menghasilkan antibodi untuk segera menghilangkan antigen, sementara sel B memori bermigrasi ke kelenjar getah bening dan menunggu kontak dengan antigen di masa mendatang.
Sel B naif hanya mengekspresikan sIgM dan sIgD pada permukaan selnya. Tetapi sel B memori mengekspresikan sIgM/sIgG/sIgA/sIgE/sIgD pada membrannya (Tabel 8.1).
Penghambatan Aktivasi sel B:
Setelah agen penginfeksi dihilangkan, inang tidak perlu lagi memproduksi antibodi.
saya. Sel plasma mati segera karena mereka memiliki masa hidup beberapa hari saja. Saat sel plasma mati, produksi antibodi lebih lanjut dihentikan.
- Tampaknya ada mekanisme umpan balik negatif yang mengatur produksi antibodi. Generasi sel plasma baru terganggu oleh penghambatan aktivasi sel B baru. Penghambatan aktivasi sel B mungkin dimediasi oleh pengikatan kompleks antigen-antibodi (terutama tipe IgG) ke sel B.
Tabel 8.1: Perbandingan sel B istirahat dan sel B memori:
|
Properti |
Sel B istirahat |
Sel B memori |
|
Permukaan |
IgM dan IgD |
IgM/IgG/IgA/ |
|
imunoglobulin |
|
IgE/IgD |
|
Reseptor komplemen |
Rendah |
Tinggi |
|
Lokasi anatomi |
Limpa, kelenjar getah bening |
Sumsum tulang, kelenjar getah bening, limpa |
|
Masa hidup |
Berumur pendek |
Berumur panjang |
Daerah Fc antibodi bergabung dengan reseptor Fc pada permukaan sel B sedangkan antigen (dalam kompleks antigen-antibodi) bergabung dengan sigs (Gambar 8.6).
sebuah†”
Pengikatan simultan kompleks antigen-antibodi ke reseptor Fc dan imunoglobulin permukaan pada sel B dapat mengganggu mekanisme pensinyalan di dalam sel B. Gangguan tersebut dapat mengakibatkan penekanan aktivasi sel B.
sebuah†”
Akibatnya, produksi sel plasma baru dan sekresi antibodi tidak terjadi.
aku aku aku. Molekul antibodi adalah protein dan terdegradasi selama periode waktu tertentu. Saat antibodi dalam sirkulasi terdegradasi, tingkat antibodi yang diinduksi terhadap antigen menurun.
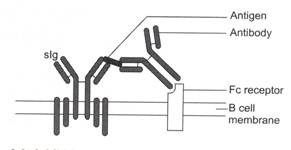
Gambar 8.6: Penghambatan aktivasi sel B.
Aktivasi sel B kemungkinan dihambat oleh pengikatan kompleks antigen-antibodi pada reseptor sig dan Fc yang terdapat pada membran sel B. Antigen yang terikat antibodi berikatan dengan slg dan wilayah Fc dari antibodi berikatan dengan reseptor Fc. Pengikatan ini mengganggu mekanisme pensinyalan di dalam sel B
Sakelar Kelas Rantai Berat:
Selalu, IgM adalah kelas antibodi pertama yang diproduksi oleh sel plasma selama respon imun primer terhadap antigen. Tetapi ketika klon sel B berkembang biak, sel anak yang mampu menghasilkan kelas imunoglobulin lain (seperti IgG atau IgA atau IgE atau IgD) muncul. Feno menon ini disebut peralihan kelas atau peralihan isotipe. Pergantian kelas terjadi dengan penataan ulang gen yang mengkode wilayah konstan imunoglobulin.
Tetapi tidak ada perubahan dalam pengkodean gen untuk wilayah variabel. Oleh karena itu, setiap kelas imunoglobulin dari klon sel B tertentu akan memiliki spesifisitas antigen yang sama, yaitu mereka akan bergabung dengan antigen yang sama. Pilihan untuk beralih ke kelas imunoglobulin baru (IgG/IgA/IgE/IgD) dipengaruhi oleh banyak faktor, seperti jaringan tempat aktivasi dan proliferasi sel B terjadi serta efek beberapa sitokin pada sel B.
saya. Lingkungan mikro di bagian usus Peyer mendukung peralihan ke kelas IgA.
- IFNγ mempromosikan peralihan kelas ke IgG1.
aku aku aku. IL-4 mempromosikan peralihan kelas ke IgE.
Respon Imun Primer dan Sekunder:
Respon imun yang diinduksi pada saat masuknya antigen pertama kali ke dalam inang disebut respon imun primer. Respon imun yang diinduksi selama masuknya antigen serupa yang kedua dan selanjutnya ke dalam inang disebut respon imun sekunder.
Respon imun primer:
Respon imun primer terhadap antigen dijelaskan dalam empat fase.
1. Lag (laten) fase:
Lag phase adalah selang waktu antara waktu masuknya antigen dengan waktu terdeteksinya antibodi terhadap antigen dalam darah. Pada manusia fase lag sekitar satu minggu. Selama fase lag, antigen diproses dan dipresentasikan ke sel T; sel B diaktifkan dan sel plasma mulai mengeluarkan antibodi.
2. Fase eksponensial:
Fase eksponensial adalah periode di mana tingkat antibodi meningkat dengan cepat. Periode ini mencerminkan jumlah besar antibodi yang disekresikan oleh sejumlah besar sel plasma.
3. Fase stabil (dataran tinggi):
Selama fase dataran tinggi, tingkat antibodi tetap pada tingkat yang konstan dalam jangka waktu yang lama. Sekresi antibodi dan degradasi antibodi terjadi dengan kecepatan yang kira-kira sama; dan karenanya tingkat antibodi tetap pada kondisi stabil.
4. Fase penurunan:
Selama fase menurun, tingkat antibodi perlahan menurun. Setelah penghilangan antigen, sel plasma baru tidak diproduksi; sel plasma yang sudah terbentuk mati dengan cepat dalam beberapa hari setelah diproduksi; dan akibatnya, antibodi baru tidak disekresikan. Degradasi molekul antibodi yang sudah disekresikan menyebabkan penurunan tingkat antibodi.
Fase lag bervariasi antara antigen yang berbeda dan periode tergantung pada banyak faktor. Bergantung pada kegigihan antigen, periode respons primer dapat hilang untuk berbagai periode, mulai dari beberapa hari hingga beberapa minggu.
Respon imun sekunder:
Berbeda dengan respon imun primer, konsentrasi puncak antibodi dicapai dalam 2-5 hari selama respon imun sekunder (Tabel 8.2). Hal ini disebabkan jumlah, kecepatan, dan intensitas aktivitas sel B memori yang lebih besar selama respon imun sekunder. Sel B memori merespons antigen lebih cepat daripada sel B naif. Jumlah sel B naif yang tersedia untuk respon imun primer terhadap antigen sedikit.
Padahal, setelah respons imun primer, jumlah sel B memori yang tersedia untuk merespons antigen sangat banyak. Jumlah sel B memori yang diaktifkan selama respons sekunder lebih banyak daripada jumlah sel B yang diaktifkan selama respons primer. Akibatnya tingkat antibodi selama respon sekunder adalah 100-1000 kali lipat lebih tinggi dari pada respon primer.

Antibodi kelas pertama yang diproduksi melawan antigen selama respons primer (oleh sel plasma yang berasal dari sel B naif teraktivasi) selalu IgM. Setelah itu kelas / kelas antibodi lain diproduksi melawan antigen. Sebagian besar sel plasma selama respon imun sekunder mengeluarkan antibodi IgG, IgA, atau IgE. Namun beberapa sel plasma juga mengeluarkan antibodi IgM selama respon imun sekunder.
Urutan peristiwa dari masuknya antigen ke dalam inang hingga waktu sekresi antibodi maksimal dalam respon imun primer (Misalnya, pada saat pertama kali masuknya antigen) membutuhkan lebih banyak waktu (sekitar 5 sampai 10 hari) bila dibandingkan dengan waktu yang diperlukan. untuk respon sekunder.
Alasan periode yang lebih lama ini untuk mencapai konsentrasi antibodi maksimum selama respons primer adalah karena waktu yang dibutuhkan untuk kejadian berikut:
sebuah. Sel T spesifik antigen dan sel B sedikit pada saat antigen pertama kali masuk ke inang. Pengikatan awal antigen dengan sel T spesifik dan sel B mungkin memakan waktu lebih lama.
- Antigen harus diproses, dan disajikan ke sel TH spesifik antigen .
- Sel TH yang teraktivasi harus berkembang biak dan menghubungi sel B spesifik antigen.
- Kemudian sel B yang diaktifkan harus berkembang biak dan menghasilkan sel plasma untuk mengeluarkan antibodi.
Aktivasi sel B oleh Antigen T-Independen:
Umumnya kontak antigen dengan imunoglobulin permukaan sel B saja tidak cukup untuk mengaktifkan sel B. Selain kontak antigen, sel B juga memerlukan bantuan dari sel T pembantu khusus antigen (CD4 + ) terdekat. Antigen semacam itu, yang membutuhkan bantuan sel T untuk mengaktifkan sel B, disebut antigen yang bergantung pada T
Namun, ada beberapa antigen yang dapat mengaktifkan sel B tanpa bantuan sel T pembantu. Antigen semacam itu disebut antigen T-independen. Ada dua jenis antigen T-independen (antigen TI-1 dan TI-2).
Antigen TI-1 pada konsentrasi tinggi menginduksi aktivasi sel B spesifik antigen dan sel B nonspesifik. Karena banyak sel B yang diaktifkan, antigen ini disebut aktivator sel B poliklonal (misalnya Lipopolisakarida dinding sel bakteri gram negatif).
Antigen ini juga merangsang makrofag untuk memproduksi IL-1 dan TNFa, yang menambah respon imun. Di sisi lain, antigen TI-2 tidak memiliki aktivitas poliklonal dan juga tidak mengaktifkan makrofag (misalnya polisakarida dinding sel bakteri).



