Baca artikel ini untuk mendapatkan informasi tentang latar belakang sejarah, jenis, bukti percobaan, karakteristik, mekanisme, dasar kimia dan dasar umum Induksi Embrio!
Pada embrio amfibi, sel-sel ektodermal dorsal di daerah mid-longitudinal berdiferensiasi membentuk lempeng saraf, hanya jika chorda-mesoderm berada di bawahnya. Chorda-mesoderm adalah lapisan yang dibentuk oleh sel-sel invaginasi dari daerah bibir blastoporal dorsal, yang membentuk atap archenteron. Mangold (1927) memilih sebagian kecil bibir blastoporal dorsal dari gastrula awal Triturus cristatus dan mencangkokkannya di tempat dekat bibir lateral blastopori gastrula inang T. taeniatus.
Sumber Gambar : archive.sciencewatch.com/inter/aut/images-aut/2009/09junYamaF1XL.jpg
Sel-sel cangkok bertambah banyak dan menyebar di dalam gastrula inang untuk membentuk korda-mesoderm tambahan di tempat ini. Chorda-mesoderm ini, selanjutnya menginduksi ektoderm gastrula inang untuk membentuk tabung saraf tambahan.
Sel cangkok itu sendiri membentuk notochord tambahan. Saat gastrula inang berkembang lebih jauh, ia tumbuh menjadi embrio ganda yang disatukan. Salah satu embrio adalah yang biasa, sedangkan yang kedua adalah yang diinduksi. Yang terakhir tidak mengembangkan kepala yang lengkap.
Eksperimen ini dengan jelas menunjukkan bahwa bibir blastoporal dorsal blastula memiliki kemampuan untuk menginduksi pembentukan lempeng saraf di ektoderm inang. Fenomena ini disebut induksi saraf. Bagian lain dari embrio juga dapat menginduksi pembentukan struktur lain. Pengaruh satu struktur dalam pembentukan struktur lain ini disebut induksi embrionik.
Faktanya, seluruh perkembangan organisme disebabkan oleh serangkaian induksi. Struktur yang menginduksi pembentukan struktur lain disebut induktor atau organisator. Zat kimia yang dipancarkan oleh induktor disebut evakuator. Jaringan tempat evakuator atau induktor bekerja disebut jaringan responsif.
Latar Belakang Sejarah Induksi Embrio:
Untuk penemuan induksi saraf, ahli embriologi Jerman, Hans Spemann dan muridnya, Hilde Mangold (1924) banyak bekerja dan untuk karyanya Spemann menerima Hadiah Nobel pada tahun 1935.
Kedua ilmuwan ini melakukan transplantasi heteroblastik tertentu antara dua spesies kadal air, yaitu Triturus cristatus dan Triturus taeniatus dan melaporkan bahwa bibir dorsal gastrula awal mereka memiliki kapasitas induksi dan organisasi ektoderm saraf dugaan untuk membentuk tabung saraf dan juga kapasitas. kebangkitan dan organisasi ektoderm, mesoderm dan endoderm untuk membentuk embrio sekunder yang lengkap.
Mereka menyebut bibir dorsal blastopori sebagai organisator utama karena merupakan yang pertama dalam rangkaian induksi dan memiliki kapasitas untuk mengatur perkembangan embrio kedua. Belakangan, organisator utama dilaporkan ada pada banyak hewan, misalnya pada katak (Daloq dan Pasteels, 1937); dalam siklostom (Yamada, 1938); pada ikan bertulang (Oppenheimer, 1936); pada burung (Waddington, 1933) dan pada kelinci (Waddington, 1934).
Pengatur primer dan induksi saraf telah dilaporkan pada chordata pra-vertebrata tertentu, seperti ascidia dan Amphioxus (Tung, Wu dan Tung, 1932). Pada tahun 1960 dan 1963 Curtis menyelidiki dan melaporkan bahwa pengatur gastrula Xenopus laevis dapat dibedakan dalam korteks bulan sabit abu-abu dari sel telur yang telah dibuahi.
Holtfreter (1945) memberikan penjelasan tentang bagaimana berbagai macam zat yang sama sekali tidak spesifik-asam organik, steroid, kaolin, biru metilen, senyawa sulfhidril, yang tidak memiliki kesamaan kecuali sifat beracun bagi neurulasi yang diproduksi sel sub-ektodermal di eksplan. Barth dan Barth (1968, 69) memberikan informasi lebih lanjut tentang sifat kimia dari induksi embrionik.
Jenis induksi embrionik:
Lovtrup (1974) mengklasifikasikan berbagai jenis induksi embrio ke dalam dua kategori dasar induksi endogen dan eksogen.
1. Induksi endogen:
Sel-sel embrio tertentu secara bertahap mengambil pola diversifikasi baru melalui induktor yang diproduksi oleh mereka secara endogen. Karena induktor ini, sel-sel ini mengalami transformasi diri atau diferensiasi diri. Contoh-contoh induksi tersebut dilaporkan dalam sel-sel Mesenchymal kutub ventral Echinoid dan dalam sel-sel kecil yang sarat kuning telur dari bibir dorsal blastopore amfibi.
2. Induksi eksogen:
Ketika beberapa agen eksternal atau sel atau jaringan dimasukkan ke dalam embrio, mereka memberikan pengaruhnya melalui proses pola diversifikasi pada sel tetangga melalui induksi kontak. Fenomena ini disebut induksi eksogen. Ini mungkin homotypic atau heterotypic tergantung pada fakta bahwa apakah induktor masing-masing memprovokasi pembentukan jenis jaringan yang sama atau berbeda (Grobstein, 1964).
Dalam induksi homotipik, sel yang berdiferensiasi menghasilkan induktor. Induktor tidak hanya berfungsi untuk mempertahankan keadaan sel yang sebenarnya, tetapi juga menginduksi sel-sel yang berdekatan untuk berdiferensiasi menurutnya, setelah melewati batas sel. Contoh terbaik dari induksi eksogen heterotipik adalah pembentukan sumbu embrionik sekunder oleh notochord presumtif yang ditanamkan pada amfibi.
Bukti eksperimental untuk induksi:
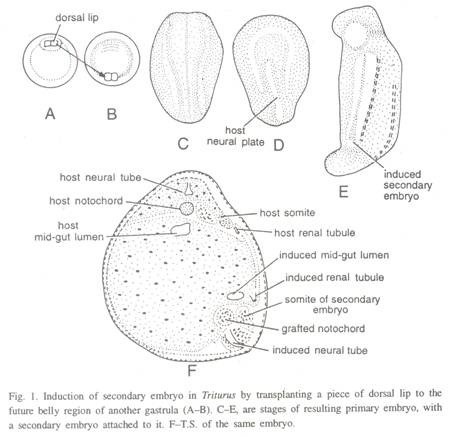
Spemann dan Mangold (1924) mentransplantasikan secara heteroplastik sepotong bibir dorsal blastopori gastrula awal kadal berpigmen, Triturus cristatus dan mencangkokkannya di dekat bibir ventral atau lateral blastopori gastrula awal kadal berpigmen T. taeniatus. Sebagian besar cangkok masuk ke dalam interior dan berkembang menjadi notochord dan somite dan menginduksi ektoderm inang untuk membentuk tabung saraf, meninggalkan potongan jaringan yang sempit di permukaan.
Dengan perkembangan embrio inang, seluruh sistem organ tambahan diinduksi pada area penempatan cangkok. Kecuali untuk bagian anterior kepala, hampir terbentuk embrio sekunder lengkap yang terdiri dari organ-organ tambahan. Bagian posterior kepala tampak seperti yang ditunjukkan oleh sepasang rudimen telinga.
Karena dalam percobaan ini jenis transplantasi yang terlibat adalah heteroplastik, ditemukan bahwa notochord embrio sekunder hanya terdiri dari sel cangkok; somit sebagian terdiri dari cangkok dan sebagian sel inang (Gbr. 1).
Beberapa sel, yang tidak berinvaginasi selama gastrulasi, tertinggal di tabung saraf. Sebagian besar tabung saraf, bagian somit, tubulus ginjal, dan dasar telinga embrio sekunder terdiri dari sel inang.
Cangkok menjadi berdiferensiasi sendiri dan pada saat yang sama menginduksi jaringan inang yang bersebelahan untuk membentuk sumsum tulang belakang dan struktur lain termasuk somit dan tubulus ginjal. Spemann (1938) menggambarkan bibir dorsal dari gastrula awal sebagai “penyelenggara utama” dari proses gastrulatif.
Namun, organisasi hasil embrio sekunder dari serangkaian interaksi induktif dan perubahan diferensiasi diri pada jaringan inang dan donor. Oleh karena itu, sekarang istilah “induksi embrionik” atau “interaksi induktif” lebih disukai. Bagian yang menjadi sumber induksi disebut “induktor”.
Ciri-ciri penyelenggara:
Organiser memiliki kemampuan untuk diferensiasi diri dan organisasi. Ia juga memiliki kekuatan untuk menginduksi perubahan di dalam sel dan mengatur sel-sel di sekitarnya, termasuk induksi dan pengaturan awal tabung saraf. Organisator primer menentukan ciri-ciri utama aksiasi dan organisasi embrio vertebrata.
Induksi adalah proses seperti alat, yang digunakan oleh pusat aktivitas ini melalui mana ia memengaruhi perubahan sel-sel di sekitarnya dan dengan demikian memengaruhi organisasi dan diferensiasi. Sel-sel di sekitarnya, yang diubah oleh proses induksi, pada gilirannya dapat bertindak sebagai pusat induktor sekunder dengan kemampuan mengatur sub-area tertentu.
Dengan demikian, transformasi blastula akhir menjadi kondisi terorganisir dari gastrula akhir tampaknya bergantung pada sejumlah induksi terpisah, semuanya terintegrasi menjadi satu kesatuan yang terkoordinasi oleh “stimulus formatif” dari organisator utama yang terletak di lempeng pre-chordal. area sel endodermal-mesodermal dan material chorda-mesodermal yang berdekatan dari gastrula awal.
Kekhususan regional penyelenggara:
Eksperimen pewarnaan vital Vogt dengan telur kadal telah menunjukkan bahwa bahan yang secara berturut-turut membentuk bibir blastoporal dorsal bergerak maju sebagai atap archenteron. Transplantasi yang diambil dari daerah ini juga dapat menginduksi embrio sekunder atau perut inang baru yaitu atap archenteron bertindak sebagai induktor utama dengan cara yang sama seperti jaringan bibir dorsal. Induksi induktor saraf ditemukan spesifik regional dan spesifisitas regional dikenakan pada organ yang diinduksi oleh induktor.
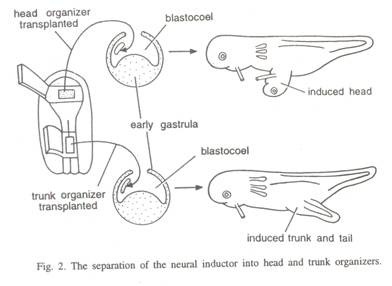
Oleh karena itu, kapasitas induktif bibir blastoporal bervariasi baik secara regional maupun temporal. Sebagian besar bahan blastoporal dorsal dan dorso-lateral diperlukan untuk cangkok untuk menginduksi embrio sekunder yang kurang lebih lengkap. Spemann (1931) menunjukkan bahwa selama gastrulasi bagian anterior dari atap archenteric berinvaginasi di atas bibir dorsal blastopore sebelumnya.
Bibir blastopori dorsal dari gastrula awal berisi organiser archenteric dan deuterocephalic dan bibir blastopori dorsal dari gastrula akhir berisi organisator spinocaudal. Induksi yang dihasilkan oleh bibir dorsal blastopori yang diambil dari gastrula awal dan akhir berbeda sesuai dengan pengecualian; yang pertama cenderung menghasilkan organ kepala dan yang kedua cenderung menghasilkan organ batang dan ekor (Gbr. 2).
Saat invaginasi berlanjut dan bibir dorsal tidak lagi terdiri dari calon endo-mesoderm kepala tetapi secara progresif menjadi calon mesoderm batang; ia bertindak sebagai induktor batang-ekor. Daerah paling kaudal dari atap arkenteron, pada kenyataannya, secara khusus menginduksi somit ekor dan mungkin jaringan mesodermal lainnya. Atap archenteron menginduksi kelas jaringan yang sama sekali berbeda; berbagai jaringan saraf dan meso-ektodermal di bagian anteriornya dan berbagai jaringan mesodermal di bagian paling posteriornya.
Oleh karena itu, perbedaan dalam kapasitas induksi spesifik ada antara tingkat kepala dan batang atap archenteron dan terkait dengan diferensiasi regional jaringan saraf menjadi archencephalic (termasuk otak depan, mata, lubang hidung), deuterencephalic (termasuk otak belakang, vesikel telinga). ) dan komponen spinocaudal. Dengan demikian, atap archenteron terdiri dari induktor kepala anterior termasuk induktor archencephalic dan induktor deuterencephalic dan batang atau induktor spinocaudal.
Induksi primer dan bulan sabit abu-abu:
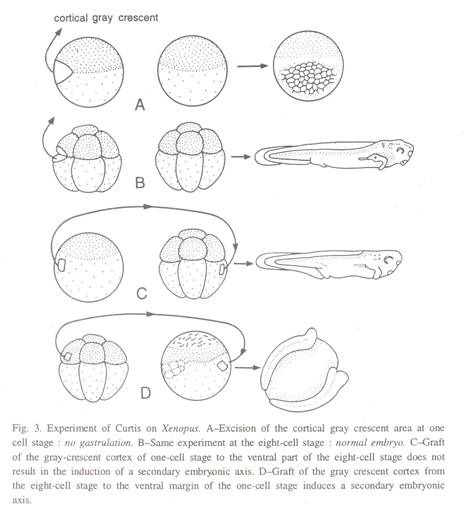
Daerah bibir dorsal blastopore pada permulaan gastrulasi dapat ditelusuri kembali ke bulan sabit abu-abu dari telur amfibi yang belum dibuahi. Dipahami oleh beberapa ahli biologi perkembangan bahwa bahan bulan sabit dari korteks telur memulai gastrulasi dan memiliki kapasitas induksi saraf. ASG Curtis (1963) melakukan serangkaian percobaan mencangkokkan bagian korteks telur yang telah dibuahi dari katak bercakar, Xenopus laevis pada awal pembelahan.
Dalam satu percobaan, korteks bulan sabit abu-abu dikeluarkan dari telur yang telah dibuahi dan diamati bahwa pembelahan sel meskipun berjalan tanpa gangguan, gastrulasi gagal terjadi (Gbr. 3Ð ). Dalam percobaan lain, korteks bulan sabit abu-abu dari telur yang belum dibuahi dipotong dan ditransplantasikan ke posisi ventral telur kedua, sehingga telur yang menerima pencangkokan memiliki dua bulan sabit abu-abu di sisi yang berlawanan.
Akibatnya, telur membelah untuk membentuk blastula, yang mengalami dua gerakan gastrulasi terpisah untuk menghasilkan dua sistem saraf primer yang terpisah, notochord dan somit terkait (Gbr. 3D). Eksperimen serupa yang dilakukan pada tahap delapan sel menunjukkan bahwa sesuatu telah terjadi selama interval pendek yang diwakili oleh tiga pembelahan pertama.
Korteks sabit abu-abu dari tahap delapan sel masih mempertahankan kapasitas induktifnya ketika dicangkokkan ke tahap yang lebih muda (Gbr. 3C). Penghapusan bulan sabit abu-abu pada tahap ini tidak lagi menghambat gastrulasi berikutnya dan perkembangan normal, sifat bulan sabit yang hilang digantikan dari daerah kortikal yang berdekatan (Gbr. 3B).
Menurut Curtis, perubahan organisasi kortikal menyebar ke seluruh permukaan telur selama pembelahan kedua dan ketiga, mulai dari bulan sabit abu-abu; ketika perubahan ini selesai, interaksi, mungkin bersifat biofisik, dapat terjadi di antara berbagai bagian korteks.
Mekanisme induksi saraf:
Perkembangan ektoderm yang menutupi atap arkenteron menjadi jaringan saraf menunjukkan aksi langsung pada sel ektodermal, baik melalui interaksi permukaan atau melalui perantaraan kimiawi.
(1) Salah satu kemungkinan yang luas adalah interaksi permukaan sel pada antarmuka induktif. Kontak dari dua lapisan selular dapat memberikan suatu alat dimana pola struktur atau geometri atau perilaku membran sel ektodermal diubah secara langsung oleh sel korda mesodermal di bawahnya.
Dengan demikian, konfigurasi spasial dari membran yang terakhir dapat menyebabkan perubahan dalam konfigurasi spasial dari membran sel ektodermal, ini pada gilirannya menghasilkan perubahan interior sel yang menentukan perkembangannya menjadi pelat saraf. Susunan morfologis semacam ini dapat menjelaskan transmisi efek induktif yang cepat dan efektif.
(2) Kemungkinan luas lainnya adalah mediasi kimia dari efek induktif. Oleh karena itu, zat kimia atau zat yang diproduksi dan dilepaskan dengan menginduksi sel korda mesoderm pada antarmuka arkenteron-ektoderm dapat bertindak atas atau memasuki sel ektodermal untuk memulai aktivitas seluler yang mengarah pada perkembangan saraf. Banyak bukti mendukung gagasan pertukaran materi antar sel dan juga menunjukkan bahwa zat yang dapat menyebar dapat bertindak sebagai stimulus induktif yang efektif.
Dasar kimia induksi saraf:
Hasil berbagai penelitian untuk menjelaskan mekanisme induksi dan untuk mengidentifikasi zat kimia atau zat yang diduga terlibat belum memberikan hasil yang baik. Ditemukan bahwa banyak jaringan yang berbeda, embrionik atau dewasa, dari berbagai spesies yang berbeda, mampu menginduksi jaringan saraf pada embrio amfibi. Selain itu, beberapa jaringan asing ditemukan menjadi induktor yang jauh lebih kuat setelah dibunuh dengan perlakuan panas atau alkohol.
Fakta ini tetap bertentangan dengan konsep ‘penyelenggara bertopeng’ yang hadir secara universal, dirilis di wilayah induktor utama. Beberapa agen anorganik seperti yodium dan kaolin, cedera lokal, paparan larutan salin dengan pH terlalu tinggi atau rendah, menyebabkan diferensiasi saraf pada ektoderm. Temuan ini menetapkan konsep besar awal pengatur embrio kimia master dari sitolisis subletal Holtfreter. Ini memiliki konsep cedera sel reversibel yang membebaskan induktor saraf.
Zat kimia yang berbeda baik bulan sabit abu-abu atau bibir dorsal atau chordamesoderm dipisahkan dengan metode biokimia yang berbeda untuk mengetahui molekul yang menyebabkan induksi saraf dan kemudian kapasitas induktif setiap molekul diuji secara terpisah. Beberapa percobaan menunjukkan bahwa evokator atau zat penginduksi adalah protein.
Upaya melelahkan dilakukan oleh ahli embriologi yang berbeda untuk memahami mekanisme sebenarnya dari induksi saraf. Beberapa teori telah dikemukakan untuk memahami mekanisme induksi saraf, di antaranya yang paling penting adalah sebagai berikut:
1. Teori denaturasi protein induksi saraf:
Menurut Ranzi (1963) induksi saraf dan pembentukan notochord berhubungan dengan denaturasi protein. Situs pembentukan notochord adalah sabit abu-abu amfibi, yang merupakan pusat aktivitas metabolisme yang tinggi. Pusat-pusat aktivitas metabolisme yang lebih besar seperti itu berhubungan dengan tempat denaturasi protein.
2. Teori gradien induksi saraf:
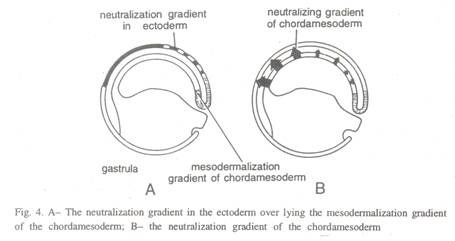
Toivonen (1968) dan Yamada (1961) menyatakan bahwa dua faktor kimia yang berbeda terlibat dalam aksi induktor primer. Dari dua faktor ini, satu adalah agen neuralizing dan yang lainnya adalah agen mesodermalizing. Eksperimen ini dilakukan dengan sumsum tulang dan hati yang didenaturasi sebagai induktor.
Spesifisitas regional sumbu embrio muncul dari interaksi antara dua gradien: prinsip penetralan memiliki konsentrasi tertinggi di sisi dorsal embrio dan berkurang secara lateral, sedangkan prinsip mesodermalisasi hadir sebagai gradien antero-posterior dengan puncaknya di daerah posterior. .
Di anterior prinsip penetralan bertindak sendiri untuk menginduksi struktur otak depan, lebih posterior prinsip mesodermalisasi bekerja bersama dengan prinsip penetralisir untuk menginduksi struktur otak tengah dan otak belakang, sementara di bagian posterior tingkat konsentrasi tinggi dari gradien mesodermal menghasilkan struktur spino-caudal (Gbr. 4).
3. Hipotesis satu faktor induksi saraf:
Nieuwkoop (1966) menggunakan notochord hidup sebagai induktor, mendalilkan bahwa hanya satu faktor yang pertama membangkitkan ektoderm untuk membentuk jaringan saraf dan kemudian menyebabkan ektoderm berubah menjadi struktur yang lebih posterior dan mesodermal (Gbr. 5) yang terlibat.
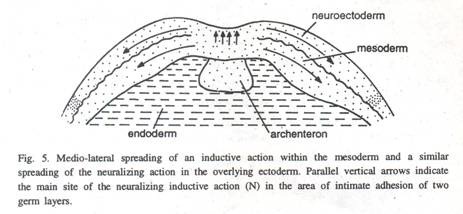
Dalam satu percobaan, yang terdiri dari menggabungkan ektoderm gastrular terisolasi dengan sepotong notochord dan kemudian menghilangkan jaringan notochord setelah jangka waktu yang bervariasi, ditemukan bahwa hanya 5 menit paparan induktor menyebabkan bagian dari ektoderm berubah menjadi struktur otak dan mata. .
4. Teori ion induksi saraf:
Menurut Barth dan Barth (1969), proses induksi yang sebenarnya dapat dimulai dengan pelepasan ion dari bentuk terikat, mewakili perubahan rasio antara terikat dengan ion bebas di dalam sel gastrula awal. Induksi sel saraf dan pigmen dalam agregat kecil calon epidermis dari gastrula katak ditemukan tergantung pada konsentrasi ion natrium.
Induksi normal sel saraf dan pigmen oleh mesoderm pada eksplan kecil dari bibir dorsal dan zona marginal lateral gastrula awal bergantung pada konsentrasi eksternal natrium. Dengan demikian, induksi embrio normal bergantung pada sumber ion endogen dan bahwa pelepasan ion intraseluler terjadi selama gastrulasi lanjut.
Dasar genik dari induksi saraf:
Ada bukti bahwa jaringan komponen induktor saraf menjadi terdiferensiasi sebelum sel ektodermal. Selama proses ini, tingkat transkripsi mRNA dan aktivasi diferensial gen menjadi berlipat ganda, sedangkan diferensiasi sel ektodermal hanya terjadi setelah pertengahan gastrulasi.
Menurut percobaan yang dilakukan oleh Tiedemann (1968), setelah 2 sampai 7 hari penanaman bibir blastopori dorsal gastrula Triturus muda dengan ektoderm yang berdekatan dalam media yang mengandung jumlah yang cukup dari Actinomycin-D untuk menghambat sintesis RNA, induksi tidak dapat terjadi, tetapi beberapa diferensiasi otot dan notochord terjadi. Ini menunjukkan bahwa mRNA dengan transkripsi dari DNA diperlukan, yang juga membutuhkan kehadiran Actinomycin-D. Oleh karena itu, tidak ada induksi saraf yang dapat dideteksi dalam percobaan ini.
Waktu induksi saraf:
Induksi saraf terjadi pada saat bahan chordamesoderm bergerak dari bibir dorsal blastopore ke dalam dan ke depan (Saxen dan Toivonen 1962). Rangsangan induktif menunjukkan gradien waktu, yang mungkin penting sehubungan dengan peristiwa aksi dan reaksi.
Induksi embrio pada chordata yang berbeda:
Meskipun induksi saraf pertama kali ditemukan pada amfibi urodele, ditemukan bahwa bibir dorsal blastopore dan atap archenteron vertebrata lain memiliki fungsi yang sama. Chordamesoderm di semua vertebrata menginduksi sistem saraf dan organ indera. Induktor saraf telah diselidiki dalam chordata berikut:
(1) Pada Cyclostomes, terutama pada lamprey, sifat induksi saraf terletak pada sel korda mesodermal dugaan bibir dorsal blastopore.
Sebelum siklostom, pada Ascidian, blastomer yang berbeda dari tahap delapan sel memiliki nasib dugaan berikut- (i) dua blastomer kutub hewan anterior menghasilkan epidermis kepala, palp, dan otak dengan dua struktur sensorik berpigmennya, (ii) dua blastomer kutub hewan posterior menghasilkan epidermis, (iii) dua blastomer vegetal anterior menghasilkan notochord, sumsum tulang belakang dan sebagian usus (iv) dua sel vegetal posterior menghasilkan mesenkim, otot dan sebagian usus.
Dari percobaan ini, Raverberi (1960) menyimpulkan bahwa pembentukan dan diferensiasi otak oleh dua blastomer hewan anterior bergantung pada induksi dua blastomer vegetal anterior, yang bertindak sebagai induktor saraf. Lebih lanjut disimpulkan bahwa dua blastomer vegetal anterior memunculkan beragam jaringan, yaitu endoderm, notochord, dan sumsum tulang belakang.
(2) Wu dan Tung (1962) membuktikan adanya penyelenggara utama dan induksi saraf di Amphioxus. Mereka mentransplantasikan potongan-potongan jaringan dari permukaan dalam bibir blastopore dorsal dari gastrula awal Amphioxus ke dalam blastocoel embrio lain pada tahap yang sama (Gbr. 6) dan mengamati bahwa embrio sekunder berkembang di daerah ventral inang dengan notochord dan mesoderm dihasilkan oleh cangkok dan tabung saraf dari jaringan inang.
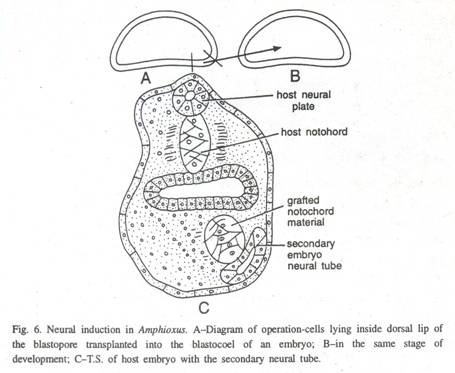
Dengan demikian, jaringan chordal Amphioxus gastrula memiliki kekuatan induksi saraf, sementara jaringan mesodermal dan endodermal memiliki sedikit kekuatan induktif.
(3) Pada ikan bertulang, induksi embrio berkembang baik sekunder dihasilkan dengan mentransplantasikan tepi posterior blastodisc yang sesuai dengan bibir dorsal blastopori, ke dalam blastocoel embrio lain (Gbr. 7) atau dengan mentransplantasikan chordamesoderm dan ektoderm. Induksi saraf juga diperoleh dengan mentransplantasikan bibir dorsal blastopore di sturgeon.
(4) Pada katak, induksi embrio sekunder dapat dihasilkan oleh bibir dorsal blastopori yang ditransplantasikan ke dalam blastocoel gastrula muda, dengan cara yang hampir sama seperti pada kadal air dan salamandar.
(5) Pada reptil archenteron memiliki aktivitas penginduksian yang sama seperti pada vertebrata lain tetapi tidak ada bukti eksperimental terjadinya induktor saraf.
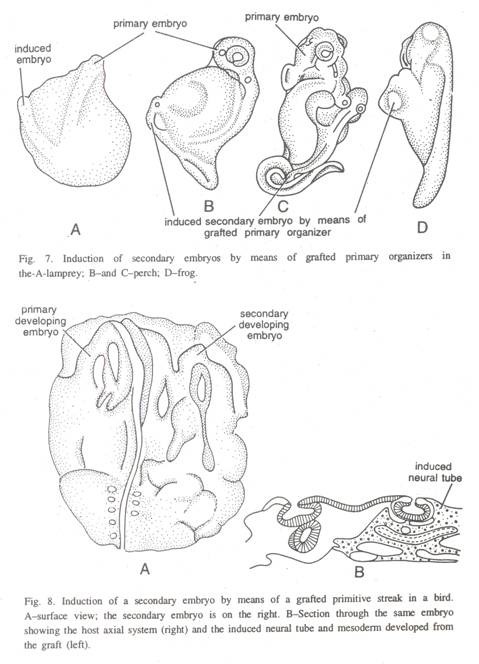
(6) Pada burung keberadaan organisator utama ditetapkan oleh Waddington dan rekan kerjanya. Separuh anterior dari garis primitif adalah bagian penginduksi yang mirip dengan bibir blastopori pada amfibi. Dalam percobaan, seluruh blastoderm dikeluarkan dari telur pada awal gastrulasi dan ditanam secara in vitro pada bekuan plasma darah.
Dari embrio lain, bagian dari garis primitif kemudian disisipkan antara epiblast dan hypoblast, induksi embrio sekunder diperoleh. Coretan primitif ditemukan bergantung pada hipoblas yang mendasari untuk pembentukannya (Gbr. 8).
(7) Induksi saraf yang berhasil dilakukan pada embrio kelinci dengan membudidayakan blastodisc awal pada bekuan plasma dan menanamkan coretan primitif anak ayam sebagai induktor. Jaringan dari gastrula mamalia ditemukan memiliki kompetensi untuk induksi saraf. Ujung anterior embrio kelinci, dengan dua pasang somit, menginduksi pelat saraf pada embrio ayam saat ditempatkan di bawah blastoderm ayam.
Jenis lain dari induksi embrionik:
Seiring dengan pertumbuhan gastrulasi, berbagai sistem organ embrio mulai berdiferensiasi dan memperoleh kekuatan untuk menginduksi diferensiasi struktur atau organ yang kemudian terbentuk seperti mata, telinga, tungkai dan paru-paru, dll. Organ-organ ini mengembangkan sifat pengorganisasian dan menjadi sumber induksi.
Oleh karena itu, rangkaian penyelenggara ini dapat disebut sebagai penyelenggara sekunder, tersier, dan kuaterner. Perkembangan progresif organ embrionik bergantung pada induksi berurutan. Satu jaringan embrionik berinteraksi dengan yang berdekatan dan mendorongnya untuk berkembang dan proses ini berlanjut secara berurutan.
Perkembangan mata:
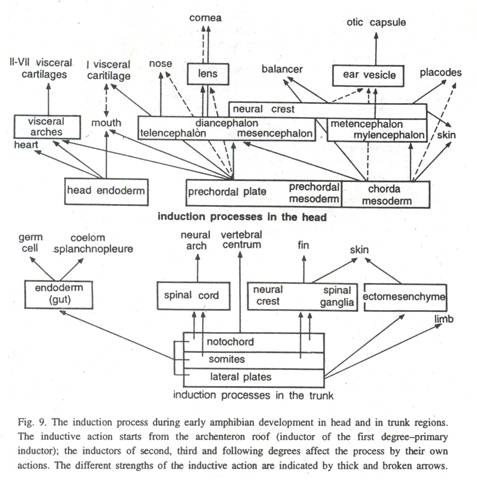
Chorda mesoderm, pengatur utama menginduksi pembentukan otak depan dan area optik di bagian anterior embrio. Daerah optik evaginates membentuk vesikel optik. Dengan invaginasi berubah menjadi struktur seperti cawan berdinding ganda, cawan optik yang bertindak sebagai pengatur sekunder untuk menginduksi pembentukan pengatur tersier untuk membentuk kornea.
Lapisan mesenkim yang tertinggal di depan bilik mata depan bergabung dengan ektoderm somatik (epidermis) di atasnya dan membentuk kornea, koroid, dan sklera (Gbr. 9). Dengan demikian seluruh proses perkembangan tampaknya menjadi penyebab induksi dan interaksi saja. Jumlah induksi sekunder atau tersier seperti alur hidung, vesikel optik, lensa, kornea dan sebagainya melibatkan reaksi ektodermal.