Kompleks Golgi: Terminologi, Kemunculan, Morfologi, Struktur, Polarisasi Struktural dan Biokimia, Komposisi Kimia, Fungsi dan Asal Usul!
Pada tahun 1898 dengan menggunakan metode pewarnaan perak, Golgi menemukan struktur retikuler di dalam sitoplasma. Nama “Aparatur Golgi†umumnya diberikan untuk struktur ini membingungkan karena menunjukkan hubungan yang pasti dengan proses fisiologis sel.
Saat ini tampaknya lebih tepat untuk menggunakan nama “substansi Golgi” atau “kompleks Golgi”, untuk merujuk pada bahan yang memiliki sifat pewarnaan khusus ini. Karena indeks biasnya mirip dengan matriks, kompleks Golgi sulit diamati pada sel hidup. Penggunaan mikroskop elektron telah memberikan gambaran yang berbeda dari komponen ini, dan struktur sub-mikroskopiknya telah terungkap.
Selama bertahun-tahun kompleks Golgi dianggap sebagai artefak dari berbagai prosedur fiksasi dan pewarnaan. Dengan kata lain, banyak ilmuwan percaya bahwa struktur yang diamati selama berbagai prosedur mikroskop dan disebut Golgi sebenarnya tidak ada dalam sel hidup. Guilliermond (1923), Parat (1927), Walker dan A lien (1921), muncul keraguan sehubungan dengan keberadaan kompleks Golgi.
Terminologi kompleks Golgi:
Holmgren menyebut kompleks Golgi sebagai trophospongium (Cajal menyebutnya sebagai kanal Golgi-Holmgren). Baker menggunakan istilah lipochondria karena dugaan kandungan lipidnya. Istilah Kompleks Dalton diberikan setelah nama pengamatnya Dalton pada tahun 1952. Sjostrand mengusulkan istilah membran-sito untuk sistem Golgi.
Sosa telah menyarankan nomenklatur berikut untuk kompleks Golgi:
- Golgiokinesis :
Divisi aparat Golgi selama divisi nuklir.
- Golgiosom :
Sel-sel yang diproduksi oleh Golgiogenesis disebut sebagai Golgiosom yang digambarkan sebagai bahan Golgi pada invertebrata.
- Golgiolisis :
Proses pembubaran aparatus Golgi.
- Gogiorrhexis :
Fragmentasi pada aparatus Golgi.
- Golegiogenesis :
Pembentukan dan diferensiasi tubuh Golgi selama perkembangan embrionik.
- Golgio-sitoarsitektur :
Studi struktur sel dalam kaitannya dengan aparatus Golgi.
Kejadian:
Kompleks Golgi terjadi di semua sel kecuali sel prokariotik (yaitu, Mycoplasma, bakteri dan ganggang hijau biru) dan sel eukariotik jamur tertentu, sel sperma bryophyta dan pteridophyta, sel tabung saringan dewasa tanaman dan sperma matang dan darah merah sel hewan.
Morfologi kompleks Golgi:
Morfologi kompleks Golgi bervariasi dari sel ke sel tergantung pada jenis sel di mana mereka ditemukan. Dua bentuk kompleks Golgi telah diamati.
- Bentuk lokal :
Pada sel vertebrata yang terpolarisasi (yang memiliki basis dan puncak), kompleks Golgi terjadi secara tunggal dan menempati posisi tetap. Itu terletak di antara nukleus dan kutub Sekretori. Hal ini paling baik terlihat pada sel tiroid, pada sel eksokrin pankreas dan sel mukosa usus.
- Bentuk menyebar :
Pada beberapa sel khusus vertebrata (sel saraf dan sel hati), pada sebagian besar sel tumbuhan dan sel invertebrata ditemukan beberapa unit kompleks Golgi tersebar bersama dengan unsur retikulum endoplasma. Setiap unit disebut dictyosome. Dalam sel hati terjadi sebanyak 50 dictyosomes per sel dan dalam sel tumbuhan tertentu jumlahnya dapat mencapai hingga ratusan.
Membentuk:
Bentuk kompleks Golgi cukup bervariasi pada berbagai jenis sel somatik hewan. Bahkan di dalam sel yang sama terdapat variasi tahapan fungsional yang berbeda. Bentuknya, bagaimanapun, konstan dengan setiap jenis sel. Ini bervariasi dalam bentuk dari massa kompak ke jaringan filamen yang menyebar.
Nomor:
Jumlah tumpukan Golgi per sel sangat bervariasi, tergantung pada jenis selnya—mulai dari satu hingga ratusan. Ada satu sel besar di beberapa sel sementara di Paramoeba ada dua sel. Dalam Stereomyxa (spesies Amoeba) terdapat banyak kompleks Golgi. Sel saraf, sel hati, dan sebagian besar sel tumbuhan juga memiliki banyak kompleks Golgi, ada sekitar 50 di sel hati.
Dalam rizoid alga terlihat lebih dari 25.000 kompleks Golgi (Sievers 1965). Kompleks Golgi bahkan dapat menjelaskan sebagian besar volume sel di beberapa sel khusus. Salah satu contohnya adalah sel goblet dari epitel usus, yang mengeluarkan lendir ke dalam usus; glikoprotein dalam lendir diglikosilasi terutama di kompleks Golgi.
Ukuran:
Ukurannya juga bervariasi. Ini besar di sel saraf dan kelenjar dan kecil di sel otot. Secara umum kompleks Golgi berkembang dengan baik saat sel dalam keadaan aktif. Ketika sel menjadi tua, kompleks tersebut semakin berkurang ukurannya dan menghilang.
Posisi:
Posisi kompleks Golgi relatif tetap untuk setiap jenis sel. Dalam sel-sel yang berasal dari ektodermal, kompleks Golgi terpolarisasi sejak masa embrio antara nukleus dan perifer (Cajal, 1914). Dalam sel eksokrin sekretori yang umumnya memiliki polarisasi khas, kompleks Golgi ditemukan antara nukleus dan kutub sekretori.
Di kelenjar endokrin, polaritas organoid ini bervariasi, kecuali di tiroid, yang berorientasi ke pusat folikel. Pada sel yang lebih muda dan seringkali pada sel yang lebih tua letaknya paling sering di satu sisi nukleus tetapi dalam kasus tertentu dapat mengelilinginya seluruhnya. Pada sel ganglion tikus posisinya perinuklear.
Struktur Rinci Kompleks Golgi:
Dalton dan Felix (1954) menggambarkan kompleks Golgi di epididimis tikus setelah mengambil mikrograf elektron pertama.
Deskripsi kompleks Golgi berikut adalah gabungan berdasarkan karya beberapa penulis:
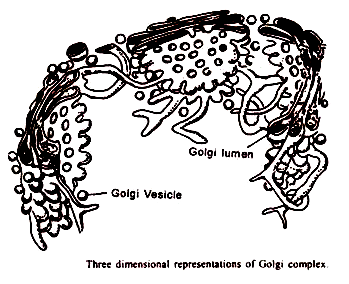
1. Sisterna:
Cisternae atau sakula mirip dengan RE permukaan halus, dan muncul di bagian sebagai tumpukan kantung yang dibatasi membran yang berjarak dekat. Jumlah sakulus bervariasi dari 4 hingga 8 di sebagian besar jenis sel hewan dan tumbuhan. Di Euglena, jumlahnya bisa mencapai 20.
Selaput kantung kira-kira setebal 60 sampai 70 A° yang membungkus rongga dengan lebar sekitar 150 A° yang ujung-ujungnya sering melebar. Menurut sebagian besar penulis, ada dua wajah cisternae yang terdefinisi dengan baik yaitu, cembung dan cekung; yang terakhir ini umumnya disebut sebagai muka dewasa atau pembentuk atau distal dan sisi cembung diasumsikan sebagai muka yang belum matang atau eksitasi atau proksimal, cisterane terletak dalam susunan paralel yang dipisahkan satu sama lain oleh ruang sekitar 200 sampai 300 A 0 .
Apa yang menyatukan mereka belum diketahui tetapi dalam beberapa sel lapisan tipis buram elektron, kadang-kadang bahan padat terlihat di antara sakulus yang pada daerah tertentu lebih menonjol yang Amos dan Grimstone (1968) menerapkan istilah node. Mollenhauer et al., (1973) mengeksplorasi secara rinci elemen intercisternal dan plak di kompleks Golgi tanaman tertentu.
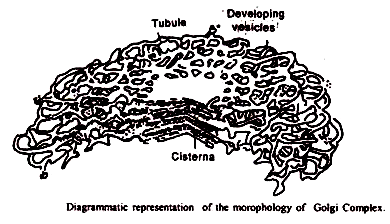
2. Tubulus:
Dari daerah perifer cisternae muncul jaringan tubulus datar yang kompleks dan beranastomosis dengan diameter 300 hingga 500 A°. Clowes dan Juniper (1969) membandingkan jaringan tubular ini dengan piringan renda.
3. Vesikel:
Vesikel adalah kantung kecil seperti tetesan yang tetap melekat pada tubulus di pinggiran cisternae.
Mereka adalah dari dua jenis berikut:
(a) Vesikel halus :
Vesikel halus berdiameter 20 hingga 80µ. Mereka mengandung bahan sekretori (sering disebut vesikel sekretori) dan bertunas dari ujung tubulus cisternal di dalam jaring. Seringkali lebih dari satu sambungan tubulus, dan mungkin mengisi, satu vesikel pembentuk.
(b) Vesikel berlapis :
Vesikel yang dilapisi adalah tonjolan bulat, berdiameter sekitar 50 µm dan dengan permukaan kasar. Mereka ditemukan di pinggiran organel, biasanya di ujung tubulus tunggal dan secara morfologis sangat berbeda dari vesikel sekretorik. Fungsi mereka tidak diketahui.
4. Vakuola Golgian:
Ini adalah kantung bulat besar yang ada di wajah Golgi yang semakin matang. Ini dibentuk baik oleh cisternae yang diperluas atau oleh fusi vesikel sekretorik. Vakuola diisi dengan beberapa zat amorf atau granular.
Kompleks Golgi secara struktural dan Biokimia Terpolarisasi:
Kompleks Golgi memiliki dua wajah yang berbeda: cis, atau wajah yang membentuk dan trans, atau wajah yang matang. Permukaan cis terkait erat dengan bagian transisi yang halus dari RE kasar. Pada sel sekretorik, muka trans adalah muka yang paling dekat dengan membran plasma: di sini, vesikel sekretorik besar ditemukan secara eksklusif berhubungan dengan muka trans tumpukan Golgi, dan membran pembentuk vesikel sekretorik seringkali bersambung dengan membran plasma. wajah trans dari cisterna terakhir (“trans-paling†).
Sebaliknya, vesikel Golgi kecil terlokalisasi lebih merata di sepanjang tumpukan, protein biasanya dianggap memasuki tumpukan Golgi dari ER di sisi cis dan keluar ke beberapa tujuan di sisi trans; namun, baik jalur pasti mereka melalui kompleks Golgi maupun bagaimana mereka melakukan perjalanan dari cisterna ke cisterna di sepanjang setiap tumpukan tidak diketahui.
Kedua wajah kompleks Golgi berbeda secara biokimia. Misalnya, variasi ketebalan membran Golgi dapat dideteksi di seluruh tumpukan dalam kasus tertentu, dengan sisi cis lebih tipis (mirip ER) dan sisi trans lebih tebal (mirip membran plasma).
Yang lebih mencolok adalah hasil yang diperoleh ketika tes histokimia tertentu digunakan bersamaan dengan mikroskop elektron untuk melokalisasi protein tertentu di dalam kompleks Golgi. Beberapa dari tes ini mengungkapkan enzim yang terikat membran, aktivitas yang menunjukkan polaritas berbeda dalam lokalisasinya di dalam tumpukan Golgi.
Temuan biokimia yang sangat menarik adalah penemuan bahwa enzim lisosom, seperti asam fosfatase, terkonsentrasi dengan cisterna paling trans dari tumpukan Golgi dan di dalam beberapa vesikel berlapis di dekatnya. Ini menunjukkan bahwa vesikel spesifik yang berangkat ke lisosom berkumpul di wilayah ini.
Protein sekretori ditemukan dengan metode histokimia di semua cisternae yang ditumpuk, meskipun vesikel sekretorik besar di mana produk ini terkonsentrasi dan hanya terkait dengan cisterna Golgi trans-paling.
Komposisi kimia:
Mengenai komposisi kimia kompleks Golgi, telah dibuktikan bahwa terdapat zat-zat berikut:
1. Fosfolipid:
Komposisi fosfolipid membran Golgi berada di antara membran endoplasma dan membran plasma.
2. Protein dan Enzim:
Kompleks Golgi dari sel tumbuhan dan hewan yang berbeda menunjukkan variasi yang besar dalam kandungan protein dan enzim. Beberapa enzim adalah ADPase, ATPase, NADPH cytochrome-C-reductase, glycosy1 transferases, galactosy1 transferase, tiamin pirofosfat dll.
3. Karbohidrat:
Sel tumbuhan dan hewan memiliki beberapa komponen karbohidrat yang sama, seperti glukosarin, galaktosa, glukosa, manosa, dan fruktosa. Tanaman Golgi kekurangan asam sialat, tetapi terjadi dalam konsentrasi tinggi di hati tikus. Beberapa karbohidrat seperti xilulosa dan arabinosa hanya ada dalam sel tumbuhan.
4. Vitamin С:
Fraksi vitamin С yang disimpan dalam kompleks Golgi telah ditunjukkan oleh Tomitte. Menurutnya kompleks Golgi menyimpan vitamin С dan melepaskannya perlahan ke dalam sitoplasma dalam jumlah yang cukup untuk mencegah oksidasi produk sel.
Fungsi kompleks Golgi:
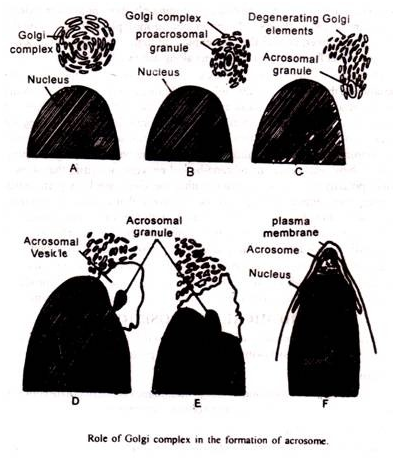
1. Pembentukan Akrosom pada Spermiogenesis:
Selama pematangan sperma kompleks Golgi berperan dalam pembentukan akrosom (Burgos dan Fawcett, 1955).
Pada tahap awal, Golgi muncul sebagai tubuh bulat, terdiri dari cisternae yang tersusun dalam tumpukan paralel dan banyak vesikel kecil. Yang belakangan selalu terjepit dari cisternae. Saat perkembangan berlangsung, kompleks Golgi menjadi tidak beraturan dan vakuola besar dibentuk oleh pelebaran kantung cisternal.
Di tengah vakuola atau vakuola besar ini terdapat butiran padat, butiran proakrosomal. Butiran yang berasal dari kompleks Golgi ini terus tumbuh di dalam vakuola melalui proses yang dikenal sebagai pertambahan. Vakuola dan granula ini mendekati kutub anterior membran nukleus, yang merupakan granula akrosom.
Dengan pemanjangan spermatid, vesikel akrosom menyebar ke permukaan nukleus dan akhirnya runtuh bersama membran nukleus, membentuk bahan penutup. Granula akrosom menjadi akrosom yang terletak di puncak nukleus dan tampaknya terdiri dari enzim-enzim tertentu yang terlibat dalam proses pembuahan.
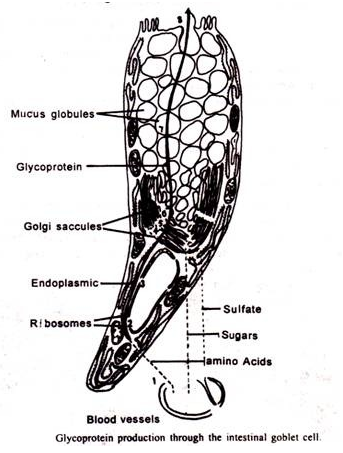
2. Sintesis dan sekresi polisakarida:
Studi tentang sel goblet dengan autoradiografi dan mikroskop elektron telah menetapkan hubungan timbal balik antara sintesis protein, penambahan karbohidrat dan sulfasi. Sel-sel piala usus besar menghasilkan mucigen. Bahan sekretorik ini mengandung sebagian besar karbohidrat.
Kompleks Golgi ditemukan tepat di atas nukleus. Menjelang permukaan bebas sel secara bertahap memperbesar butiran mucigen. Cisternae proksimal dari kompleks Golgi tidak menunjukkan adanya pembengkakan, tetapi pada jarak tertentu di seberang tumpukan, cisternae distal tiba-tiba berubah menjadi butiran mucigen. Cisternae distal terus berubah menjadi butiran mucigen setiap 2-4 menit. Cisternae proksimal baru terbentuk sebagai kompensasi.
3. Peran dalam sekresi:
Kompleks Golgi dianggap memainkan beberapa peran dalam fungsi sekretori sel. Tetapi pertanyaannya adalah bahwa mereka mengeluarkan atau mensintesis beberapa zat sendiri atau mereka hanyalah gudang di mana produk sekretori yang disekresikan di tempat lain di dalam sel, disimpan dan dipekatkan begitu saja.
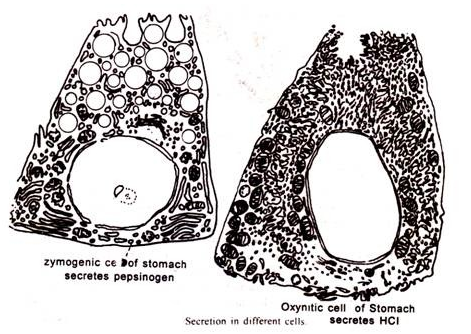
Dari studi Palade et al. 1962 siklus sekretori ini sekarang terdefinisi dengan baik dan mencakup empat langkah dalam kasus sel asinar pankreas dan mereka adalah:
(i) Penggabungan asam amino ke dalam protein pada permukaan retikulum endoplasma kasar.
(ii) Pemindahan protein sekretorik yang baru lahir ini ke dalam cisternae retikulum endoplasma kasar.
(iii) Transportasi intraseluler dari protein ini ke kompleks Golgi.
(iv) Migrasi butiran zymogen menuju puncak sel di mana mereka dibuang ke lumen.
4. Peran badan Golgi dalam oogenesis:
Srivastava (1965) telah memberikan tinjauan singkat tentang kompleks Golgi selama oogenesis. Menurut Afzelius (1956), kompleks Golgi telur bulu babi, seperti yang terlihat di bawah mikroskop elektron, terdiri dari tumpukan lamellae yang membentuk dinding kantong datar, yang kadang-kadang membengkak.
Ada beberapa indikasi pembagian melintang dari badan-badan ini. Sotelo (1959) dan Sotelo dan Porter (1959) telah menggambarkan kompleks Golgi pada tikus-ovum seperti yang terlihat di bawah mikroskop elektron dan menemukan lokalisasi nuklir juxtra dari organel ini pada oosit awal.
Pada tahap selanjutnya, ini terpecah menjadi fragmen dan pada tahap ketiga, ini bergerak menuju korteks. Dalam semua kasus ini, struktur mereka tetap berupa susunan ramping, profil ganda (kantung pipih) dan vesikel bulat.
Pada oosit awal, kompleks tersusun secara kompak. Pada tahap selanjutnya, kumpulan profil yang terpisah, dikelilingi oleh vesikel kecil ditemukan tersebar di zona kortikal. Pada oosit awal, kompleks Golgi dan sentrosom terkait erat.
5. Penyerapan senyawa:
Hirsch et al., telah menemukan bahwa ketika gula besi diumpankan ke hewan, besi diserap di kompleks Golgi (Kedrowsky). Van Teel telah menunjukkan bahwa sistem Golgi juga menyerap senyawa tembaga dan emas. Kedrowsky telah menunjukkan bahwa kompleks Golgi Opalina dapat menyerap bismutose (senyawa albumin dan bismut) dan protargol (senyawa albumin dan perak). Dengan demikian, Kirkman dan Severinghaus menyatakan bahwa kompleks Golgi bertindak sebagai membran kondensasi untuk pemusatan produk menjadi tetesan atau butiran.
6. Pembentukan dinding sel tumbuhan:
Dinding sel tanaman terdiri dari fibril yang sebagian besar mengandung polisakarida, bersama dengan beberapa lipid dan protein. Selama sitokinesis pelat sel terbentuk di antara dua inti anak, dan di sekelilingnya terdapat membran yang kemudian menjadi membran plasma sel anak. Ada bukti jelas bahwa polisakarida terbentuk di kompleks Golgi dan dipindahkan ke dinding sel baru yang diletakkan saat sel masih tumbuh.
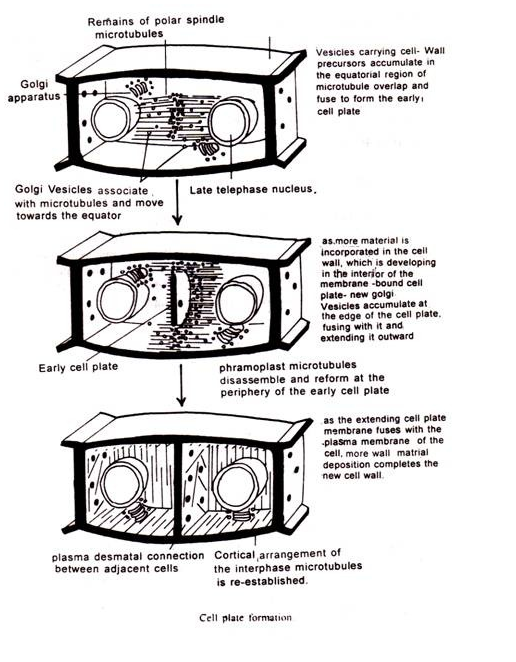
Zat seperti pektin dan hemiselulosa, yang membentuk matriks pelat sel yang memisahkan membran plasma, juga disumbangkan oleh kompleks Golgi.
7. Pembentukan kristal intraseluler:
Dalam isopod laut, Limnoria lingmorum, yang merupakan bentuk liang terdapat midglands yang sel-selnya terdiri dari kristal. Panjangnya berkisar hingga 30 A° dan tebal 15 A°. Telah dibuktikan bahwa kristal-kristal tersebut dibentuk oleh kompleks Golgi dan diketahui mengandung protein dan zat besi. Mereka tanpa membran penutup dan biasanya berbentuk bulat. Mereka prihatin dengan aktivitas sekretori.
8. Pembentukan tetesan Protein Susu:
Di kelenjar susu mencit diproduksi tetesan protein yang terkait dengan kompleks Golgi. Tetesan ini biasanya terbuka ke permukaan sel melalui fusi membran penutupnya dengan membran plasma.
9. Pembentukan lisosom dan vakuola:
Lisosom primer terbentuk dari membran Golgi dengan cara yang sama seperti vesikel sekretorik. Ada bukti bagus bahwa diktiosom mengakumulasi enzim hidrolitik di daerahnya yang lebih matang. Beberapa vakuola dalam sel tumbuhan ditemukan mengandung sejumlah kecil enzim hidrolitik dan diduga berasal dari kompleks Golgi.
10. Pembentukan pigmen:
Pada banyak sel tumor dan kanker mamalia, kompleks Golgi digambarkan sebagai tempat asal butiran pigmen (melanin).
11. Pengaturan keseimbangan cairan:
Homologi telah disarankan antara kompleks Golgi dan vakuola kontraktil dari Metazoa dan Protozoa yang lebih rendah. Vakuola kontraktil mengeluarkan kelebihan air dari sel. Dalam Protozoa tertentu kompleks Golgi juga berkaitan dengan pengaturan keseimbangan cairan.
Asal Kompleks Golgi:
Tiga sumber berbeda telah diusulkan dari mana kompleks Golgi baru dapat muncul:
1. Dari retikulum endoplasma:
Essner dan Novikоff (1962) dan Balok dan Kessel (1968) telah mengusulkan bahwa cisternae Golgi muncul dari ER. Retikulum endoplasma kasar setelah mensintesis protein spesifik kehilangan ribosom dan berubah menjadi RE halus. Vesikel sementara kecil mencubit dari ER halus. Ini bermigrasi ke dictyosome.
Saat mencapai wajah pembentuk dictyosome, sekering ini membentuk cisternae baru dan dengan demikian berkontribusi pada pertumbuhannya. Dengan peleburan vesikel-vesikel ini, cisternae baru terbentuk terus-menerus pada permukaan pembentuk dan pada permukaan yang matang, cisternae lama pecah menjadi vesikel sekretorik. Jadi Golgi menunjukkan fenomena aliran membran.’
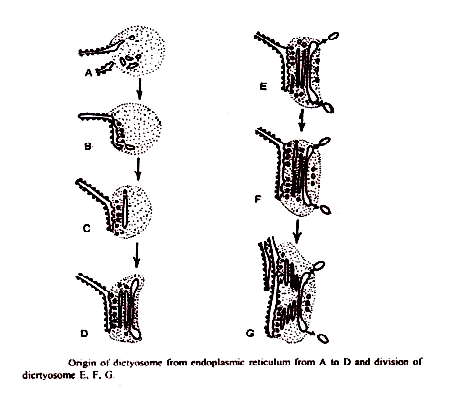 2. Dari membran inti:
2. Dari membran inti:
Bouch (1965) menjelaskan asal-usul Golgi dari membran luar selubung inti pada ganggang coklat. Vesikel terjepit dari membran inti luar yang menyatu membentuk cisternae pada permukaan pembentuk dictyosome.
Adanya zona eksklusi dalam kaitannya dengan RE halus atau membran inti, terjadinya zona eksklusi pada benih dorman tanaman tingkat tinggi dan pembentukan diktiosom dari zona ini pada benih berkecambah memberikan bukti yang mendukung dua teori di atas tentang asal usul dictyosome.
3. Dengan pembagian dictyosome yang sudah ada sebelumnya:
Telah diamati bahwa selama pembelahan sel pada tumbuhan dan hewan, jumlah diktiosom meningkat dan jumlah diktiosom di setiap sel anak segera setelah pembelahan hampir sama dengan jumlah di sel induk sebelum pembelahan, dari kaleng dan langsung lainnya. Pengamatan pada sel-sel yang membelah diduga bahwa diktiosom juga membelah selama pembelahan sel.



