Kromosom: morfologi, struktur, heteropiknosis, pita kromosom, dan ultrastruktur kromosom!
Kromosom pertama kali terlihat oleh Hofmeister (1848) dalam sel induk serbuk sari Tradescantia dalam bentuk tubuh yang diwarnai gelap. Istilah kromosom (Gr: chrom=warna; soma=badan) digunakan oleh Waldeyer (1888) untuk menunjukkan afinitasnya yang besar terhadap zat warna dasar.
Signifikansi fungsional mereka dijelaskan oleh IV.S. Sutton (1900) ketika ia menelusuri kesejajaran antara segregasi kromosom selama meiosis dan transmisi faktor herediter selama gametoeenesis. Tinjauan umum tentang morfologi kromosom telah dipublikasikan oleh Heitz (1935), Kuwada (1939), Geitler (1940) dan Kaufmann (1948).
Kromosom adalah komponen sel yang paling signifikan, terutama terlihat selama mitosis dan meiosis. Kehadiran mereka telah ditunjukkan jauh sebelum mereka dinamai “kromosom” oleh Waldeyer pada tahun 1888.
Sebuah kromo beberapa dapat dianggap sebagai komponen nuklir yang memiliki organisasi khusus, individualitas dan fungsi. Ia mampu bereproduksi sendiri sambil mempertahankan sifat morfologis dan fisiologisnya melalui pembelahan sel yang berurutan.
Morfologi:
Morfologi kromosom dapat menjadi studi terbaik pada metafase atau anafase mitosis ketika mereka hadir sebagai organel tertentu, yang paling padat atau melingkar.
Nomor:
Jumlah kromosom pada spesies tertentu biasanya konstan yang mengandung jumlah kromosom diploid (2n) dalam sel somatiknya dan jumlah kromosom haploid (gamet atau tereduksi) (n) dalam sel kelaminnya (sperma dan ovum). Jumlah kromosom bervariasi dari satu hingga beberapa ratus di antara spesies yang berbeda
Misalnya pada Ascaris megalocephala berjumlah 2, sedangkan pada protozoa tertentu (Aggreta) terdapat lebih dari 300 kromosom, pada Paramecium 30 sampai 40, pada radiolaria sebanyak 1600, pada Hydra vulgaris 32, Musca domestica 12, Rana esculenta 26 , Columba livia 80, Oryctolagus cuniculus 44, Gorilla gorilla 48 dan Homo sapiens (manusia) 46.
Jumlah kromosom juga berguna untuk taksonomi. Pada angiospermae jumlah haploid yang paling sering adalah 12 dan anggota kelompok ini berkisar antara 3 hingga 16. Demikian pula pada jamur, jumlah haploid berkisar antara 3 hingga 8.
Pada primata jumlah haploid ini adalah dari 16 hingga 30. Kumpulan kromosom haploid ini terdapat pada inti gamet sedangkan pada sel diploid akan terdapat dua genom. Sel diploid adalah sel somatik dalam tubuh. Sel-sel diploid mendapatkan set kromosom diploid dengan penyatuan gamet jantan dan betina haploid dalam reproduksi seksual.


Ukuran:
Kromosom berkisar, rata-rata dari 0,5 sampai sekitar 30µ panjangnya dan dari 0,2 sampai Зµ dengan diameter. Jumlah relatif kromosom umumnya berbeda dalam nukleus tetapi pada suatu waktu semua kromosom sel mungkin berukuran sama. Sel tumbuhan biasanya memiliki kromosom yang lebih besar daripada sel hewan.
Trillium memiliki kromosom yang dapat mencapai panjang 32µ pada metafase. Tanaman monokotil biasanya memiliki kromosom yang lebih besar daripada dikotil yang mengandung lebih banyak kromosom. Di antara hewan, belalang, jangkrik, belalang sembah, kadal air, dan salamander memiliki kromosom besar.
Variasi ukuran kromosom dapat diinduksi oleh sejumlah agen lingkungan:
- Sel yang membelah pada suhu rendah memiliki kromosom yang lebih pendek dan lebih padat daripada sel yang membelah pada suhu tinggi.
- Colchicine adalah alkaloid yang mengganggu pembentukan spindel dan pembelahan sel. Ini cenderung memendekkan kromosom.
- Pembelahan yang cepat dan berulang cenderung menghasilkan kromosom yang lebih kecil. Tampaknya laju pembelahan sel berlangsung lebih cepat daripada pembentukan bahan kromatin seperti biasa.
- Pada tumbuhan, jumlah fosfat dalam media nutrisi berpengaruh nyata pada ukuran kromosom; konsentrasi tinggi memberikan kromosom yang lebih besar daripada tanaman yang kekurangan fosfat. Karena fosfat merupakan bagian integral dari molekul asam nukleat, tampaknya jumlah asam nukleat dalam kromosom dapat bervariasi untuk memberikan perubahan ukuran.
Membentuk:
Bentuk kromosom dapat berubah dari fase ke fase dalam proses pertumbuhan sel dan pembelahan sel yang berkelanjutan. Pada fase istirahat atau tahap interfase sel, kromosom terjadi dalam bentuk struktur tipis, melingkar, elastis dan kontraktil, seperti benang yang dapat diwarnai, benang kromatin.
Pada metafase dan anafase, kromosom menjadi tebal dan berserabut. Setiap kromosom mengandung zona bening, yang dikenal sebagai sentromer atau kinetokor, sepanjang panjangnya. Sentromer membagi kromosom menjadi dua bagian, setiap bagian disebut lengan kromosom.
Posisi sentromer bervariasi dari kromosom ke kromosom dan memberikan bentuk yang berbeda ke yang terakhir sebagai berikut:
- Telosentris :
Kromosom mirip batang yang memiliki sentromer di ujung proksimal dikenal sebagai kromosom telosentris.
- Akrosentrik :
Kromosom akrosentrik berbentuk seperti J tetapi memiliki sentromer di salah satu ujungnya dan dengan demikian memberikan lengan yang sangat pendek dan lengan yang sangat panjang. Belalang (Acrididae) memiliki kromosom akrosentrik.
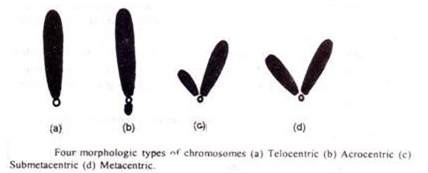
- Sub-metasentrik :
Kromosom sub-metasentrik berbentuk L. Dalam hal ini, sentromer terjadi di dekat pusat atau di bagian tengah kromosom dan dengan demikian membentuk dua lengan yang tidak sama.
- Metasentrik :
Kromosom metasentrik berbentuk V dan pada kromosom ini sentromer terjadi di tengah dan membentuk dua lengan yang sama. Amfibi memiliki kromosom metasentrik.
Struktur Kromosom:
Deskripsi mikroskopis cahaya sebelumnya kromosom, atau kromatid, dianggap terdiri dari benang melingkar yang disebut kromonema yang terletak di matriks. Kromosom seharusnya ditutupi oleh pelikel membran.
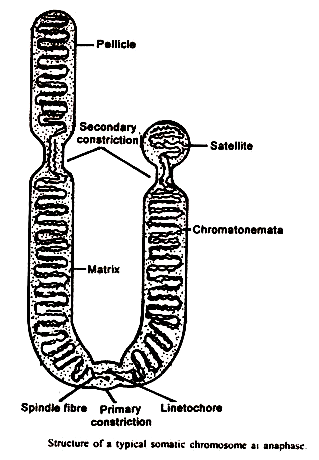
Studi mikroskop elektron kemudian menunjukkan bahwa tidak ada pelikel membran yang pasti mengelilingi kromosom. Struktur lain yang ada dalam kromosom termasuk kromatid, sentromer, penyempitan sekunder, pengatur nukleolus, telomer, dan satelit seperti yang diberikan di bawah judul berikut:
Kromatid:
Selama metafase kromosom tampaknya memiliki dua benang yang disebut kromatid, yang terjalin dalam matriks kromosom. Kedua kromatid ini disatukan pada satu titik sepanjang panjangnya di daerah penyempitan kromosom.
Kromatid-kromatid ini benar-benar adalah kromonemata melingkar spiral (sing., kromonema) pada metafase. Filamen melingkar pertama kali diamati oleh Baranetzky, pada tahun 1880, dalam sel induk serbuk sari Tradescantia dan disebut chromonema oleh Vejdovsky pada tahun 1912.
Kromonema dapat terdiri dari 2, 4 atau lebih fibril tergantung pada spesiesnya. Jumlah fibril dalam kromonema ini mungkin bergantung pada fase yang berbeda karena pada satu fase mungkin mengandung satu fibril dan fase lainnya mungkin mengandung dua atau empat fibril. Serat-serat kromonema ini saling melingkar.
Gulungan terdiri dari dua jenis:
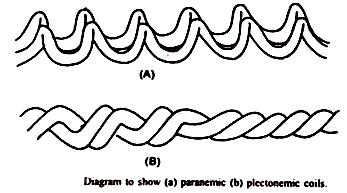
- Kumparan paranemik :
Ketika fibril kromonemal mudah dipisahkan satu sama lain, gulungan semacam itu disebut gulungan paranemik.
- Kumparan pletonemik:
Di sini fibril kromonemal terjalin erat dan tidak dapat dipisahkan dengan mudah. Kumparan semacam itu disebut kumparan plectonemik. Tingkat lilitan fibril kromonemal selama pembelahan sel tergantung pada panjang kromosom.
Ada tiga jenis kumparan:
(i) Kumparan utama chromonema memiliki 10-30 pilin.
(ii) Kumparan kecil dari chromonema tegak lurus dengan gulungan utama dan memiliki banyak pilinan seperti yang diamati pada kromosom meiosis. Jika belum terjadi pembelahan pada tahap ini akan terjadi satu chromonema, jika sudah terjadi akan terjadi dua chromonemata.
(iii) Gulungan standar atau somatik ditemukan dalam kromonema mitosis di mana kromonemata memiliki struktur heliks, menyerupai gulungan utama kromosom meiosis.
kromomer:
Kromonema kromosom tipis profase mitosis dan meiosis telah ditemukan mengandung daerah tebal dan tipis bergantian dan dengan demikian memberikan tampilan kalung di mana beberapa manik-manik terjadi pada sebuah tali.
Struktur kromonema yang tebal atau seperti manik-manik dikenal sebagai kromomer dan daerah tipis di antara kromomer disebut sebagai interkromomer. Posisi kromomer dalam kromonema ditemukan konstan untuk kromosom tertentu.
Para ahli sitologi telah memberikan berbagai interpretasi tentang kromomer. Beberapa menganggap kromomer sebagai bahan nukleo-protein terkondensasi, sementara yang lain mendalilkan bahwa kromomer adalah daerah kumparan super-dikenakan.
Pandangan selanjutnya telah dikonfirmasi oleh pengamatan mikroskop elektron. Untuk waktu yang lama, sebagian besar ahli genetika menganggap kromomer ini sebagai gen, yaitu unit keturunan.
sentromer:
Ini adalah bagian tak terpisahkan dari kromosom dan membentuk penyempitan utama pada metafase. Tanpa kromosom sentromer tidak dapat mengarahkan diri dengan baik pada pelat metafase. Karena sentromer menempati posisi konstan, maka sentromer bertanggung jawab atas bentuk kromosom.
Dengan demikian, bentuk kromosom ditentukan oleh penyempitan primer, yang terletak di titik pertemuan lengan kromosom. Di dalam penyempitan primer, ada zona bening, memiliki butiran atau sferula kecil. Wilayah bening ini dikenal sebagai sentromer (Gr. meros, part) atau kinetokor atau kinetomere.
Fungsinya dalam hal pergerakan. Ini bertanggung jawab untuk pembentukan serat kromosom di gelendong. Struktur sentromer berbentuk bulat telur, tidak ternoda, dengan diameter besar, seperti pada jagung atau mungkin seperti butiran atau sferula kecil, seperti pada Tradescantia.
Di sentromer, mungkin ada satu atau lebih butiran atau sferula kecil, yang disebut kromomer dan serat gelendong. Biasanya, setiap kromosom hanya memiliki satu sentromer. Dalam kasus seperti itu, kromosom disebut monosentris. Mungkin ada dua yaitu, disentrik atau lebih polisentrik atau dengan sentromer tersebar, seperti yang ditemukan di Ascaris megctlocephalus dan Hemiptera.
Setelah penelitian terbaru, diketahui bahwa sentromer terdiri dari tiga zona yang hadir dalam rangkap dua. Zona tengah mempertahankan hubungan kromosom dengan gelendong. Diagram yang diberikan di bawah ini menunjukkan dua saudara kromatid yang membentuk setiap kromosom metafase yang dipegang oleh suatu daerah yang memiliki siklus pembelahan khusus.
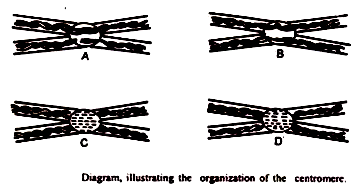
Sentromer dianggap secara fungsional terbagi ke sumbu longitudinal kromosom pada awal anafase. Pergerakannya menuju kutub diatur oleh keterikatannya pada spindel. Kadang-kadang, pembelahan juga terjadi pada sudut kanan sumbu longitudinal membentuk dua sentromer segmen, yang melekat pada dua kromatid dari masing-masing lengan.
Struktur ini, terdiri dari dua lengan, dikenal sebagai kromosom; nama itu disarankan oleh Darlingtion pada tahun 1939. Mc. Clintock (1932) telah melaporkan bahwa kerusakan tersebut juga dimungkinkan oleh sinar-x. Dalam kasus seperti itu, setiap bagian fragmen sentromer berfungsi.
Diketahui juga bahwa sentromer adalah struktur majemuk, yang bagian-bagiannya terkoordinasi dalam pembagian dan pergerakan. Sachrader (1936) dan Darlington (1939) telah menyarankan bahwa sentromer dapat dianggap homolog dengan sentriol baik secara struktural maupun teoritis.
Penyempitan Sekunder:
Selain penyempitan primer atau sentromer, lengan-lengan kromosom dapat menunjukkan satu atau lebih penyempitan sekunder (disebut penyempitan sekunder-II). Ini berbeda dari pengatur nukleolus (disebut penyempitan sekunder I), meskipun beberapa ahli sitologi juga menyebut pengatur nukleolus sebagai penyempitan sekunder.
Lokasi penyempitan II sekunder adalah konstan untuk kromosom tertentu, dan karena itu berguna untuk identifikasi kromosom. Telah dikemukakan bahwa penyempitan sekunder mewakili situs kerusakan dan fusi selanjutnya. Pada pria penyempitan sekunder II ditemukan pada lengan panjang kromosom 1, 10, 13, 16 dan Y Nucleolar Organizer (penyempitan sekunder 1).
Penyelenggara Nukleolus (Konstriksi Sekunder I):
Biasanya di setiap set kromosom diploid, dua kromosom homolog memiliki ‘penyempitan’ tambahan yang disebut pengatur nukleolar. Ini disebut demikian karena mereka diperlukan untuk pembentukan nukleolus.
Nukleolus terbentuk pada fase rekonstruksi pasca-mitosis. Di bawah mikroskop cahaya pengatur nukleolar muncul sebagai ‘penyempitan’ di dekat salah satu ujung kromosom. Bagian kromosom di luar pengatur nukleolar sangat pendek dan tampak seperti bola (satelit). Pada manusia kromosom 13, 14, 15, 21 22 dan Y memiliki pengatur nukleolar dan satelit. Kromosom yang membawa satelit disebut SA T-kromosom.
Awalan SAT adalah singkatan dari ‘Sine Acid Thymonucleionico’ (tanpa asam timonukleat atau DNA), karena kromosom pada pewarnaan menunjukkan defisiensi relatif DNA di wilayah pengatur nukleolar. Setidaknya ada dua kromosom SAT di setiap nukleus diploid.
Telomer:
Ujung kromosom bertindak berbeda dari bagian interstisial. Jika ujung atau telomer putus baik secara spontan atau dengan induksi, biasanya hilang dari nukleus pada pembelahan sel berikutnya karena kekurangan sentromer.
Ujung yang patah dari kromosom bebas yang tersisa bersifat stabil dan dapat menyatu dengan ujung kromosom lain yang patah di sekitarnya. Namun, ujung yang putus tidak akan menyatu dengan ujung yang normal. Dalam profase meiosis, telomer terkadang tertarik ke sentriol dan terlihat bermigrasi ke membran inti di dekat sentriol. Perilaku ini menghasilkan apa yang disebut sebagai tahap Buket.
Marix dari Kromosom:
Seperti yang diduga oleh beberapa ahli sitologi bahwa kromonemata tertanam dalam matriks kromatik yang dibatasi oleh pelikel. Namun, pengamatan terbaru dari studi mikroskop elektron mengungkapkan tidak adanya pelikel. Matriks tidak didefinisikan sebagai massa utama kromosom, yang secara khas Feulgen-positif. Itu dapat dihilangkan dengan cara enzimatik meninggalkan kromosom residu Feulgen-negatif.
Heteropiknosis:
Secara umum telah diamati selama berbagai tahap mitosis bahwa kromosom atau bagian tertentu dari kromosom tidak seperti itu, tetapi lebih padat daripada kariotipe lainnya. Ini mengacu pada heteropiknosis. Fenomena ini mengakibatkan penggumpalan kromosom selama pembelahan sel. Heteropycnosis mungkin positif, diikuti dengan kondensasi berlebih atau negatif, menunjukkan kondensasi di bawah atau tanpa kondensasi.
Juga telah diamati bahwa bagian tertentu dari kromosom atau seluruh kromosom mungkin tidak menunjukkan kondensasi atau heteropiknosis di semua fase. Karena heteropiknosis adalah kekhasan heterokromatin, ini membantu dalam membedakannya dari eukromatin. Ini jauh lebih lazim dalam kromosom seks, meskipun yang lain juga menunjukkannya.
Eukromatin dan Heterokromatin:
Meskipun selama interfase kromatin kromosom menyebar dalam bentuk benang-benang halus linin tetapi pada daerah tertentu kromatin dikenal sebagai daerah heterokromatin atau heterokromatin.
Heterokromatin terdiri dari dua jenis:
- Heterokromatin fakultatif, dan
- Heterokromatin konstitutif.
- Heterokromatin fakultatif :
Ini mewakili keadaan sementara inaktivasi kromatin di mana satu kromosom pasangan menjadi sebagian atau seluruhnya heterokromatik. Sebagai contoh, pada mamalia salah satu dari dua kromosom X pada sel somatik wanita menjadi heterokromatik dan membentuk badan kromatin seks atau Barr (Bar dan Bertram, 1944). Pada sel somatik laki-laki, hanya terdapat satu kromosom X dan tetap euchromatic (tidak ada badan Barr).
- Heterokromatin konstitutif:
Jenis heterochromatin menyajikan fitur yang lebih permanen dan ditemukan di kedua kromosom pasangan. Ini sangat sering ditemukan di daerah sentromerik, telomer, di daerah pengatur nukleolus atau sebagai pita di daerah kromosom lainnya. Ini terkait erat dengan nukleolus pada tumbuhan dan hewan.
Banding kromosom:
TC Hsu dan lain-lain (1969) memperkenalkan metode baru untuk pewarnaan kromosom dimana pola yang berbeda dari pita bernoda dan antar pita bernoda ringan menjadi jelas. Metode pewarnaan ini sangat penting karena memungkinkan setiap kromosom diidentifikasi secara unik, bahkan jika morfologi keseluruhannya identik. Perbedaan sekarang dapat dibuat di antara kromosom kelompok Ð yang relatif mirip. Misalnya, sekarang kita dapat mengatakan kromosom I atau 2 atau 3 alih-alih hanya sebuah kromosom grup Ð dari sistem Denver.
Berikut ini adalah metode untuk banding kromosom:
1.G-Banding:
Metode banding kromosom yang paling berguna adalah G-banding. Teknik ini dikembangkan oleh Hsu dan Arrighi. Diamati bahwa ketika kromosom diinkubasi dalam air liur diwarnai dengan pewarna Giemsa atau diolah dengan urea atau deterjen. G-band muncul di area yang kaya akan protein S. Preparat pewarnaan Giemsa lebih permanen dan membutuhkan optik dan pencahayaan mikroskop biasa.
2. Q-Banding:
Teknik ini dikembangkan oleh Casperson. Diamati, ketika kromosom diwarnai dengan mustard quinacrine dan diamati melalui mikroskop fluoresensi, daerah kromosom yang kaya akan adenin dan timin ternoda secara intens.
Daerah guanin-sitosin tetap tidak ternoda. Daerah ini disebut Q-band. Cacat pewarnaan ini adalah, noda memudar setelah beberapa saat, terlebih lagi, optik mikroskopis khusus ditambah iluminasi ultraviolet diperlukan untuk melihat pita ini.

3. C. Banding:
Teknik ini dikembangkan oleh Pardue dan Gall. Kromosom diperlakukan dengan natrium hidroksida yang kuat diikuti dengan salin hangat dan kemudian diwarnai dengan pewarnaan Giemsa. С-band terutama terlihat di sekitar sentromer dan kromosom lain yang mengandung sejumlah besar heterokromatin konstitutif yang sangat berulang.
4.R.Banding:
Pita ini muncul saat kromosom diinkubasi dalam buffer pada suhu tinggi dan diwarnai dengan pewarnaan Giemsa. Pita-R sesuai dengan daerah pada charom9osom yang memiliki protein yang kekurangan belerang. Ini adalah timbal balik dari G band.
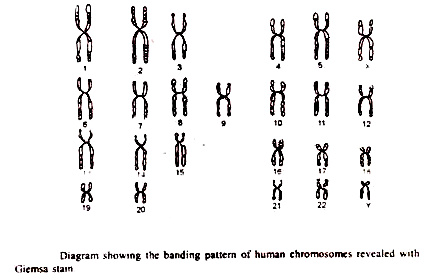
Teknik pembengkokan pewarnaan kromosom sangat berguna untuk mengetahui berbagai jenis aberasi kromosom seperti delesi, duplikasi, inversi atau translokasi. Kepastian yang lebih besar dalam mengidentifikasi seluruh kromosom atau bagian dari kromosom dengan G-band sering memungkinkan peneliti untuk mengetahui dengan tepat kromosom mana yang ada dan bagian kromosom mana yang telah mengalami penataan ulang struktural. Banding juga menyediakan sarana untuk membandingkan kariotipe spesies terkait dan untuk menggambarkan perbedaan yang tampaknya memiliki dasar evolusi.
Ultra-struktur Kromosom:
Dua pandangan telah diusulkan untuk ultra-struktur kromosom:
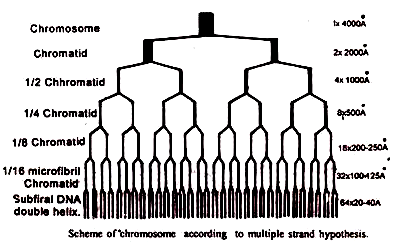
(a) Tampilan multistranded:
Hal ini dikemukakan oleh Ris (1966). Dengan mikroskop elektron unit terkecil yang terlihat dari kromosom adalah fibril dengan ketebalan 100 A°. Fibril ini mengandung dua molekul heliks ganda DNA yang dipisahkan oleh ruang 25A° melintang dan protein terkait.
Unit terbesar berikutnya adalah setengah kromatid. Setengah kromatid terdiri dari empat fibril 100 A 0 sehingga tebalnya 400 A° dan mengandung delapan heliks ganda pada DNA dan protein terkait. Dua setengah kromatid dari kromatid lengkap yang terdiri dari 16 molekul heliks DNA ganda.
Karena kromosom terdiri dari dua kromatid, maka jumlah total heliks akan menjadi 32 dan diameter 1600 A° sebelum duplikasi atau sintesis. Setelah duplikasi kromosom memiliki 64 heliks ganda DNA dengan diameter yang sesuai 3200 A°. Jumlah heliks DNA di setiap unit di atas tingkat fibril bervariasi menurut spesies. Singkatnya, kromosom terdiri dari banyak fibril mikro, yang terkecil adalah molekul nukleoprotein tunggal.
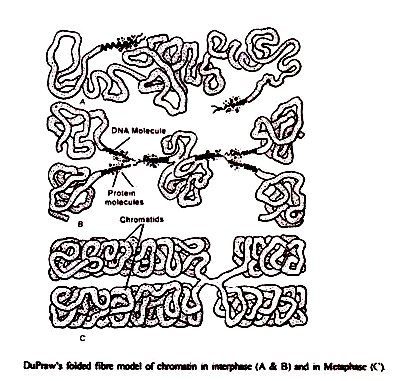
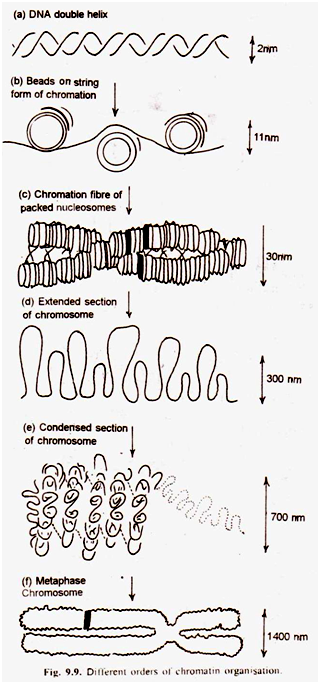
(b) Model Fibril Lipat:
DuPraw (1965) mempresentasikan model ini untuk struktur kromosme yang halus. Menurut model ini, sebuah kromosom terdiri dari satu rantai panjang DNA dan protein yang membentuk apa yang disebut fibril. Fibril dilipat berkali-kali dan terjalin secara tidak teratur untuk membentuk kromatid. Ini mengukur ketebalan 250-300 A.
Sub-unit nukleosom kromatin:
Kromatin menginformasikan unit berulang, yang disebut nukleosom. Istilah tersebut diberikan oleh Oudet et al, (1975). Nukleosom terdiri dari DNA dan protein histon. Protein membentuk partikel inti yang merupakan dua molekul Oktober dari masing-masing empat protein histon yaitu. H2a H2b, H3 dan H4. Permukaan partikel inti dikelilingi 1,75 lilitan DNA (200 pasang basa).
DNA yang menghubungkan partikel inti disebut DNA penghubung. Protein histon lain, HI terikat pada DNA penghubung. (Kornberg dan Thomas, 1974). Partikel inti berukuran tinggi 40 A° dan diameter 80 A°. Seluruh nukleosom berukuran tinggi 55 A° dan diameter 110 A 0 .
Kromosom Politen:
Balbiani pada tahun 1881 adalah orang pertama yang mengamati kromosom kelenjar ludah di kelenjar ludah larva Chironomus. Jenis kromosom raksasa ini terbatas pada jenis jaringan somatik tertentu pada serangga yang termasuk dalam ordo Diptera.
Biasanya mereka mencapai ukuran terbesar mereka di inti bulat dari kelenjar ludah larva, tetapi inti serupa sering ada di jaringan lain seperti sel-sel lapisan usus dan turunannya, tubulus Malpighi serta otot dan sel lemak dll. Dengan demikian istilah kromosom “polytene†Koller lebih disukai daripada istilah umum kromosom kelenjar ludah.
Struktur:
Struktur kromosom kelenjar ludah sangat menarik secara sitogenetik. Di sepanjang kromosom ada serangkaian pita gelap yang bergantian menjadi zona bening lainnya yang disebut interband. Pita gelap sangat ternoda dan Feulgen positif. Selanjutnya, mereka menyerap sinar ultraviolet pada 600 A°. Pita ini dapat dianggap sebagai cakram, karena menempati seluruh diameter kromosom.
Ukurannya bervariasi. Pita yang lebih panjang memiliki struktur yang lebih rumit. Mereka sering membentuk doublet, dua pita yang terletak bersebelahan dan dengan ketebalan dan bentuk yang identik. Interband adalah aspek fibrilar, tidak diwarnai dengan pewarna dasar, negatif Feulgen dan menyerap sedikit sinar ultraviolet. Selain itu, mereka menghadirkan elastisitas yang lebih besar daripada daerah pita. Keteguhan dalam situasi dan distribusi cakram atau pita dalam dua kromosom homolog (berpasangan) patut diperhatikan.
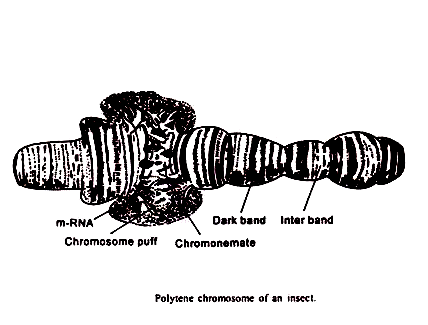
Dalam kasus Drosophila melanogaster, kromosom dari masing-masing nukleus polytene, ketika diratakan, muncul sebagai untaian halus panjang dan satu yang cukup pendek melekat pada massa pusat yang dikenal sebagai pusat kromo, yang juga melekat pada satu nukleolus besar. Hubungan antara untaian ini adalah kromosom cahaya dari set mitosis biasa dari spesies ini pada awalnya tidak jelas.
Penjelasannya tergantung pada dua fakta:
(1) Dua anggota dari setiap pasang kromosom menyatu erat sepanjang panjangnya;
(2) Sentromer dari semua kromosom bersama dengan segmen heterokromatik yang berdekatan semuanya bergabung bersama untuk membentuk pusat kromo.
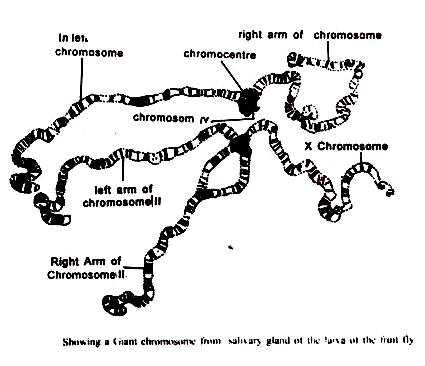
Jadi, dari enam helai, yang pendek mewakili dua kromosom IV yang menyatu, dan yang lebih panjang mewakili kromosom X, sedangkan empat sisanya adalah kaki dari kromosom kedua dan ketiga yang berbentuk ‘V’. Pada inti kelenjar air liur dari larva betina, untaian yang mewakili ‘X’ adalah ganda, seperti yang lainnya, sedangkan pada inti dari individu jantan itu tunggal. V cukup kecil dan hampir seluruhnya termasuk dalam pusat kromo.
Pusat kromo terjadi pada semua spesies Drosophila dan ukurannya tergantung pada apakah segmen heterokromatik proksimal luas atau tidak. Pada beberapa kelompok Diptera lainnya, pusat kromo famili Sciadoceridae dan Chironomidae tidak ada.
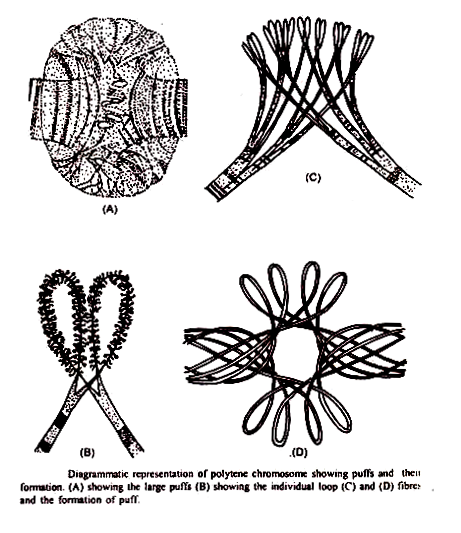
Aktivitas Kanan dan Gen Puffis dan Balbiani :
Kekhasan morfologi terpenting dari kromosom polytene adalah adanya pita dan antar pita. Brewer, Pavan, Beermann, McChelke dan lain-lain telah menemukan bahwa pada tahap-tahap tertentu perkembangan larva beberapa pita khusus dari kromosom polytene menunjukkan pembesaran.
Pita yang membesar ini dianggap sebagai unit terakhir dari hereditas-gen yang sedang bekerja. Gen-gen aktif ini berupa gumpalan-gumpalan yang tersebar di sana-sini di sepanjang kromosom kelenjar ludah. Beermann dan Clever (1964) telah menemukan bahwa kepulan menghasilkan RNA dan RNA yang dibuat dalam satu kepulan berbeda dari RNA kepulan lainnya.
Pengamatan tiupan telah menunjukkan pola aktivitas gen pada beberapa serangga yang sedang berkembang. Juga diamati bahwa hormon tertentu dan zat lain dapat memulai, menghentikan, dan mencegah beberapa aktivitas ini. Struktur halus pita individu dapat berbeda sehubungan dengan tiupan yang berada di satu lokasi pada kromosom di satu jaringan dan di lokasi lain pada kromosom yang sama di waktu lain atau di jaringan lain. Modifikasi terlokalisasi dalam struktur kromosom berbagai Diptera ini telah dicatat bertahun-tahun sebelumnya tetapi kemungkinan signifikansinya diabaikan.
Koherensi filamen kromosom dilonggarkan di daerah kembung. Cincin longgar selalu dimulai pada satu pita. Dalam tiupan kecil, pita tertentu kehilangan konturnya yang tajam dan menampilkan tampilan yang menyebar dan tidak fokus di mikroskop. Di lokus lain atau di lain waktu, pita mungkin terlihat seolah-olah telah “terbuka” menjadi cincin atau lingkaran besar di sekitar kromosom.
Struktur mirip kacang seperti itu disebut cincin Balbiani, diambil dari nama EG Balbiani, yang pertama kali mendeskripsikannya pada tahun 1881; engah dianggap karena terungkap atau uncoiling kromosom individu dalam sebuah band. Pada pengamatan bahwa jaringan spesifik dan tahapan perkembangan dicirikan oleh pola tiupan yang pasti, Beermann (1952) mendalilkan urutan tiupan tertentu mewakili pola aktivitas permainan yang sesuai. Dari aktivasi gen diferensial memang terjadi, orang dapat memprediksi bahwa gen dalam jenis sel tertentu akan mengembang secara teratur sedangkan gen yang sama di jaringan lain tidak akan menggembung.
Gen dengan sifat yang sama telah dideskripsikan dalam kelompok empat sel kelenjar ludah Chironomus. Chironomus pallidivittatus menghasilkan sekresi granular. Spesies terkait erat Chironomus tentatus mengeluarkan sekresi non-granular yang jelas dari sel yang sama.
Dalam hibrida dari kedua spesies ini, sifat ini mengikuti hukum hereditas sederhana Mendel. Beermann dan Clever (1964) mampu melokalisasi perbedaan dalam kelompok yang kurang dari 10 pita di salah satu kromosom Chironomus dan kromosom tersebut ditetapkan sebagai kromosom IV.
Sel penghasil granul C. pallidivittatus memiliki kepulan yang diasosiasikan dengan kelompok pita ini, kepulan yang sama sekali tidak ada di lokus kromosom IV yang sesuai pada Chironomus tentatus. Pada hibrida, kepulan muncul hanya pada kromosom yang berasal dari induk C pallidivittatus; hibrida menghasilkan jumlah butiran yang jauh lebih kecil daripada induknya.
Selain itu, ukuran kepulan berkorelasi positif dengan jumlah butiran. Ini mengungkapkan dengan jelas hubungan antara kepulan dan produk seluler (spesifik). Studi ini menunjukkan hubungan spesifik antara puffgene dan fungsi spesifik sel.
Teori mengenai struktur kromosom polytene:
Ada tiga teori untuk menjelaskan struktur kromosom polytene.
Dari mereka, penjelasan ketiga sebenarnya adalah kombinasi dari dua teori pertama:
- Kromosom polytene adalah hasil dari beberapa siklus reproduksi kromosom intraseluler dan terdiri dari kumpulan kromosom biasa yang terlipat. Ini adalah teori polytene yang disponsori oleh Her-twig (1935), Cooper (1938) Painter (1939) dan Beerrnann (1952).
- Kromosom polytene adalah kromosom berpasangan yang memiliki panjang dan lebar yang sangat besar dengan penambahan atau penggabungan bahan tambahan yang tidak ada pada kromosom biasa. Ini adalah konsep alveolar awal dari Metz (193 5) dan dikemukakan oleh Kodani (1942) dan Darlington (1949).
- Kromosom polytene terdiri dari bundel chromonema, ukurannya sebagian disebabkan oleh akumulasi bahan tambahan di pusat kromosom, dan atau pertumbuhan aktual panjang chromonema (Koltzof, 1934; Painter, 1934; Calvin et al, 1940; Ris dan Course, 1954; White, 1945)
Teori Politena:
Painter (1941) percaya bahwa peningkatan diameter disebabkan oleh pembesaran dan mungkin oleh duplikasi terus-menerus dari masing-masing kromomer tetapi tanpa pemisahan variabel kromonemata individu. Jadi dalam perkembangannya masing-masing kromomer asli menjadi terpecahkan dengan pemisahan melalui peregangan menjadi sejumlah kromomer yang lebih kecil.
Duplikasi pembesaran dan agregasi kromomer homolog menghasilkan penampilan pita kromatik melintang. Kromosom, oleh karena itu, menjadi multistranded sebagai polytene tetapi chromonemata individu, yang menurut Painter mungkin sebanyak 1024, sedangkan Beermann (1952) memperkirakan tingkat polytene setinggi 16.000 kali.
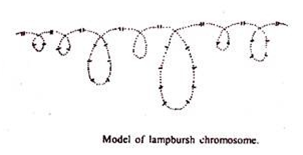
Kromosom sikat lampu:
Di seluruh kelompok hewan vertebrata, kromosom somatik menunjukkan struktur yang biasa, tetapi di dalam oosit yang sedang berkembang dari vertebrata yang memiliki telur kuning telur dan selama tahap diploten meiosis, kromosom yang sama mengalami perubahan yang luar biasa, terutama mencirikan у peningkatan panjang yang sangat besar dan penampilan rambut memancar; atau s loop yang tampak mengatur diri mereka sendiri dari penampilan seperti sikat kromo mere selama profase meiosis.
Jenis kromosom ini pertama kali dideskripsikan oleh Flemming (1882) dan Rukert (1892) memberi nama terkenal, “Lampbrush,†Ris (1951) menemukan kromosom serupa pada hiu, burung, amfibi dll. Kadang-kadang kromosom ini mencapai maksimum. berukuran hingga 800 hingga 1.000µ per kromosom.
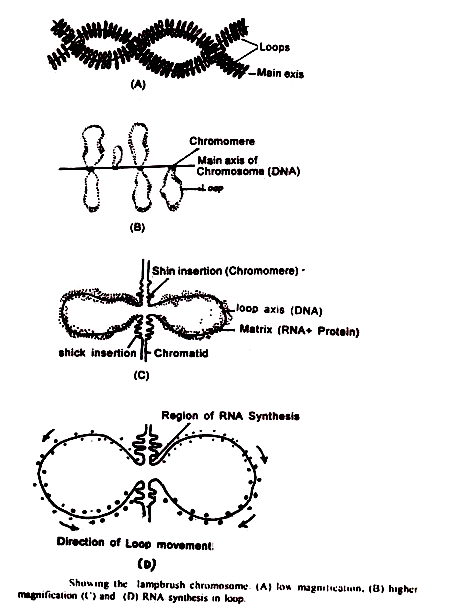
Kromosom sikat lampu memiliki sumbu kromosom pusat yang memproyeksikan serangkaian loop lateral. Loop tampaknya memproyeksikan keluar dari wilayah padat. Menurut Duryee, setiap kromosom menyerupai satu silinder plastik di mana, pada lokus tertentu, tertanam butiran kromatin.
Dalam kromosom bivalen terdapat sekitar 150-200 butiran berpasangan. Butiran-butiran ini berukuran о dua ukuran, yaitu kromiol yang lebih kecil dan kromiol yang lebih besar, yang kemudian berbentuk ellipsoidal seolah-olah terjepit di dalam matriks.

Loop lateral menimbulkan penampilan seperti sikat. Loop dianggap bahan kromatin disintesis untuk pemanfaatan eksternal, dan bukan merupakan bagian integral dari chromonemata diperpanjang dalam bentuk kumparan utama seperti yang diusulkan oleh Ris (1945).
Hipotesis Duryee (1941) tentang sintesis lateral didukung oleh fakta bahwa peregangan kromosom dengan manipulasi mikro atau kontraksi oleh ion kalsium tidak menyebabkan loop menghilang atau tergeser dan bahwa pembubaran loop oleh berbagai zat menjadi butiran tidak mempengaruhi integritas kromonemata.
Gall (1956) menunjukkan dengan cukup meyakinkan melalui mikroskop elektron bahwa loop adalah bagian dari chromonemata dan hilangnya mereka yang tampak disebabkan oleh fakta bahwa sebelum kontak mereka melepaskan lapisan asam nukleatnya.
Kromosom memiliki elastisitas yang luar biasa. Loop lateral yang memanjang dari kromomer lebih rapuh. Gall (1958) telah menafsirkan bahwa pembentukan loop adalah perubahan fisiologis reversibel yang mungkin bersifat non-genetik.
Namun, ia menunjukkan bahwa loop menunjukkan variasi dalam morfologi mereka yang juga menunjukkan, di sisi lain, bahwa mungkin setiap pasangan loop, merupakan lokus genetik yang berbeda yang bertanggung jawab untuk pembentukan beberapa produk sel tertentu.
Kromosom lampu-sikat berisi poros utama pusat yang terus menerus fleksibel. Sumbu loop dikelilingi oleh protein yang dikombinasikan dengan RNA. Mungkin loop terutama berkaitan dengan sintesis RNA, protein dan bahan kuning telur.
Untuk menjelaskan ukuran besar lingkaran sikat lampu, Callan dan Loved (1960) mendalilkan bahwa masing-masing tidak terdiri dari gen, tetapi dari sejumlah salinan duplikat, yang disusun secara linier, dari satu gen. Kromomer , unit dasar organisasi dalam kromosom sikat lampu ada dalam dua bentuk. Ada salinan ‘master’ dari gen tertentu dalam kromomer yang mirip dengan salinan gen ‘budak’ yang identik.
Dengan cara ini loop hanya berisi jumlah salinan duplikat. Gen diisolasi oleh garis ganda dan garis melintang tunggal yang menunjukkan ujung salinan duplikat. Gall dan Callan mengamati bahwa loop lateral selalu memiliki satu ujung yang tipis dan satu ujung yang lebih tebal pada titik penyisipan di kromomer.
Dipercaya juga bahwa loop yang berputar keluar dari kromomer bersatu kembali dengannya di ujung yang tebal, yang menunjukkan akumulasi RNA yang berat. Callan lebih lanjut menyarankan bahwa hanya salinan ‘budak’ yang mengambil bagian dalam sintesis RNA. Ini memastikan kemungkinan untuk mensintesis asam ribonukleat dalam jumlah besar.
Pembentukan nukleolus pada kromosom lampbrush menunjukkan pola yang tidak biasa. Mungkin ada beberapa ratus nukleolus yang mengambang bebas di dalam nukleoplasma. Signifikansi tidak dipahami dengan baik, tetapi diduga bahwa mereka harus mensintesis bahan untuk pertumbuhan.
Kromosom Aksesori atau Supernumerary:
Inti beberapa hewan dan tumbuhan memiliki, selain kromosom normal, satu atau lebih kromosom aksesori atau supernumerary. Wilson (1905) adalah ahli sitologi pertama yang mengamatinya pada serangga hemipteran, Metapodius. Sejak itu, mereka telah dilaporkan di beberapa serangga dan di banyak tumbuhan tingkat tinggi juga.
Dalam beberapa kasus, sifat dan asal mereka pasti diketahui. Namun, nenek moyang mereka belum sepenuhnya diketahui. Kromosom supernumerary biasanya berukuran lebih kecil dari jenisnya. Dianggap bahwa mereka melakukan beberapa fungsi yang belum dirusak, yang terlalu lemah untuk dideteksi secara genetik.



