Kromosom: Struktur, Fungsi, dan detail lain tentang Kromosom!
Kromosom (Gr., chrom=warna, soma= tubuh) adalah tubuh berbentuk batang, berwarna gelap yang terlihat selama tahap metafase mitosis ketika sel diwarnai dengan pewarna dasar yang sesuai dan dilihat di bawah mikroskop cahaya. Kromosom pertama kali dijelaskan oleh Strasburger (1815), dan istilah ‘kromosom’ pertama kali digunakan oleh Waldeyer pada tahun 1888.
Studi selanjutnya dengan jelas menunjukkan bahwa kromosom terdiri dari benang kromatin tipis yang disebut kromonemata yang mengalami penggulungan dan penggulungan super selama profase sehingga kromosom menjadi lebih tebal dan lebih kecil secara progresif, dan mudah diamati di bawah mikroskop cahaya.
Nomor kromosom:
Jumlah kromosom konstan untuk spesies tertentu. Oleh karena itu, ini sangat penting dalam penentuan filogeni dan taksonomi spesies. Jumlah atau set kromosom sel gamet seperti sperma dan ovum dikenal sebagai set kromosom gamet, tereduksi atau haploid.
Kumpulan kromosom haploid juga dikenal sebagai genom. Sel somatik atau tubuh sebagian besar organisme mengandung dua set haploid atau genom dan dikenal sebagai sel diploid. Sel-sel diploid mencapai set kromosom diploid dengan penyatuan gamet jantan dan betina haploid dalam reproduksi seksual.
Jumlah kromosom pada setiap sel somatik sama untuk semua anggota spesies tertentu. Organisme dengan jumlah kromosom terendah adalah nematoda, Ascaris megalocephalus univalens yang hanya memiliki dua kromosom dalam sel somatik (2n = 2).
Pada protozoa radiolaria Aulacantha ditemukan jumlah diploid kurang lebih 1600 kromosom. Di antara tanaman, jumlah kromosom bervariasi dari 2n = 4 di Haplopappus gracilis (Compositae) hingga 2n=> 1200 di beberapa pteridophyta. Jumlah kromosom beberapa hewan dan tumbuhan umum diberikan di bawah ini:
|
Hewan |
Nomor kromosom |
|
1. Paramecium aurelia |
30 – 40 |
|
2. Hydra vulgaris |
32 |
|
3. Ascaris lumbricoides |
24 |
|
4. Musca dcmestica |
12 |
|
5. Honio sapiens |
46 |
|
Tanaman |
Nomor kromosom |
|
1. Mucor heimalis |
2 |
|
2. Allium cepa |
16 |
|
3. Aspergillus nidulans |
16 |
Autosom dan kromosom seks:
Dalam sel diploid ada dua dari setiap jenis kromosom yang disebut sebagai kromosom homolog, kecuali kromosom seks. Misalnya, pada manusia, ada 23 pasang kromosom homolog (yaitu, 2n = 46).
Laki-laki manusia memiliki 44 kromosom non-seks, disebut autosom dan sepasang kromosom seks heteromorfik atau berbeda secara morfologis, yaitu satu kromosom X dan satu kromosom Y. Perempuan manusia memiliki 44 kromosom non-seks (autosom) dan sepasang kromosom seks homomorfik (mirip secara morfologis) yang ditetapkan sebagai XX.
Morfologi:
Perubahan morfologi kromosom dengan tahap pembelahan sel, dan kromosom metafase mitosis paling cocok untuk studi tentang morfologi kromosom. Pada kromosom metafase mitosis, ciri struktural berikut (kecuali kromomer) dapat dilihat di bawah mikroskop cahaya:
(1) Kromatid,
(2) Kromonema,
(3) Kromomer,
(4) Sentromer
(5) Penyempitan sekunder atau pengatur Nukleolar,
(6) Telomer dan
(7) Satelit.
Struktur dan wilayah yang dikenali dalam kromosom:
Secara struktural, setiap kromosom dibedakan menjadi tiga bagian—
(a) Pelikel,
(b) Matriks
(c) Kromonemata.
(a) Pelikel:
Ini adalah selubung luar di sekitar substansi kromosom. Ini sangat tipis dan terbentuk dari zat achromatic. Ilmuwan tertentu Darlington (1935) dan Ris (1940) menyangkal keberadaannya.
(b) Matriks:
Ini adalah substansi dasar kromosom yang mengandung kromonemata. Itu juga terbentuk dari bahan nongenik.
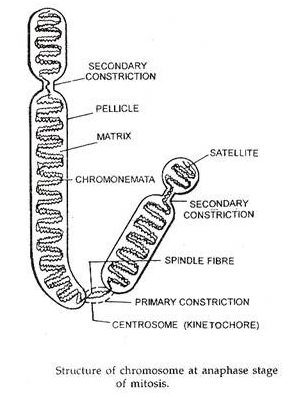
(c) Kromonemata:
Tertanam dalam matriks setiap kromosom adalah dua benang identik yang digulung secara spiral, chromonemata. Kedua chromonemata juga digulung rapat sehingga tampak sebagai utas tunggal dengan ketebalan sekitar 800A. Setiap kromonemata terdiri dari sekitar 8 mikrofibril , yang masing-masing terbentuk dari heliks ganda DNA.
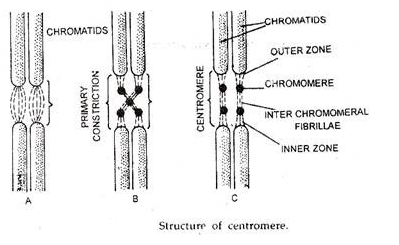
kromomer:
Dalam persiapan yang menguntungkan, kromomer dalam bentuk massa padat kecil diamati secara berkala pada kromonemata. Ini lebih berbeda pada tahap profase ketika kromonemata kurang melingkar dan paling jelas terlihat selama tahap leptoten dan zigoten dari profase meiosis.
Bagian tipis dan bernoda ringan di antara kromosom yang berdekatan disebut sebagai antar-kromomer. Posisi kromomer pada kromonemata adalah konstan untuk kromosom tertentu.
Saat berpasangan selama zigoten profase meiosis, kromosom homolog memasangkan kromomer ke kromomer. Kromomer adalah daerah DNA yang terlipat rapat dan diyakini sesuai dengan unit fungsi genetik dalam kromosom.
Kromatid:
Pada metafase mitosis setiap kromosom terdiri dari dua struktur simetris yang disebut kromatid. Setiap kromatid mengandung satu molekul DNA. Kedua kromatid terikat satu sama lain hanya oleh sentromer dan dipisahkan pada awal anafase, ketika kromatid saudara dari kromosom bermigrasi ke kutub yang berlawanan.
sentromer:
Bagian dari kromosom diakui sebagai permanen. Ini adalah struktur kecil di chromonema dan ditandai dengan penyempitan. Pada titik ini kedua chromonemata bergabung bersama. Ini dikenal sebagai sentromer atau kinetokor atau penyempitan primer. Posisinya konstan untuk jenis kromosom tertentu dan membentuk fitur identifikasi.
Pada bagian mikroskopis elektron tipis, kinetokor menunjukkan struktur trilaminar, yaitu lapisan protein luar padat setebal 10 nm, lapisan tengah dengan kerapatan rendah dan lapisan dalam padat yang terikat erat ke sentromer.
Kromosom melekat pada serat gelendong di wilayah ini selama pembelahan sel. Bagian dari kromosom yang terletak di kedua sisi sentromer merupakan lengan yang mungkin sama atau tidak sama tergantung pada posisi sentromer.
Bergantung pada jumlah sentromer, kromosom dapat berupa:
- Monosentris dengan satu sentromer.
- Disentrik dengan dua sentromer.
- Polisentrik dengan sentromer lebih dari dua seperti pada Luzula
- Asentrik tanpa sentromer. Kromosom semacam itu mewakili segmen kromosom yang baru saja rusak yang tidak bertahan lama.
- Difusi atau non-lokasi dengan sentromer tidak jelas menyebar sepanjang kromosom.
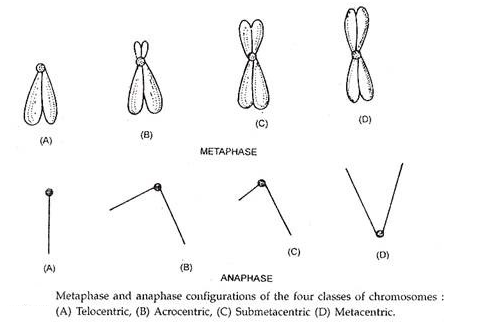
Bergantung pada lokasi sentromer, kromosom dikategorikan menjadi:
- Telosentrik adalah kromosom berbentuk batang dengan sentromer menempati posisi terminal, sehingga kromosom hanya memiliki satu lengan.
- Akrosentrik juga merupakan kromosom berbentuk batang dengan sentromer menempati posisi sub-terminal. Satu lengan sangat panjang dan yang lainnya sangat pendek.
- Kromosom submetasentrik dengan sentromer sedikit menjauhi titik tengah sehingga kedua lengan tidak sama.
- Metasentrik adalah kromosom berbentuk V dimana sentromer terletak di tengah kromosom sehingga kedua lengan hampir sama.
Sentromer mengontrol orientasi dan pergerakan kromosom pada gelendong. Ini adalah titik di mana gaya diberikan ketika kromosom bergerak menjauh selama anafase.
Penyempitan Sekunder atau Penyelenggara Nukleolus:
Kromosom selain memiliki penyempitan primer atau sentromer juga memiliki penyempitan sekunder di setiap titik kromosom. Konstan dalam posisi dan luasnya, penyempitan ini berguna dalam mengidentifikasi kromosom tertentu dalam satu set.
Penyempitan sekunder dapat dibedakan dari penyempitan primer atau sentromer, karena kromosom membengkok hanya pada posisi sentromer selama anafase. Wilayah kromosom distal ke penyempitan sekunder yaitu, wilayah antara penyempitan sekunder dan telomer terdekat dikenal sebagai satelit.
Oleh karena itu, kromosom yang mengalami penyempitan sekunder disebut kromosom satelit atau kromosom sat. Jumlah kromosom sat dalam genom bervariasi dari satu spesies ke spesies lainnya.
Nukleolus selalu dikaitkan dengan penyempitan sekunder kromosom sat. Oleh karena itu, penyempitan sekunder disebut juga nucleolus organizer region (NOR) dan kromosom sat sering disebut sebagai kromosom nucleolus organisator. NOR dari setiap sat-kromosom mengandung beberapa ratus salinan gen yang mengkode RNA ribosom (rRNA).
Telomer:
Ini adalah ujung khusus dari kromosom yang menunjukkan diferensiasi dan polaritas fisiologis. Setiap ekstremitas kromosom karena polaritasnya mencegah segmen kromosom lain menyatu dengannya. Ujung kromosom dikenal sebagai telomere. Jika kromosom putus, ujung yang putus dapat menyatu satu sama lain karena kurangnya telomere.
Kariotipe dan Idiogram:
Sekelompok tumbuhan dan hewan yang terdiri dari suatu spesies dicirikan oleh seperangkat kromosom, yang memiliki ciri-ciri konstan tertentu seperti jumlah kromosom, ukuran dan bentuk kromosom individu. Istilah kariotipe telah diberikan kepada kelompok karakteristik yang mengidentifikasi satu set kromosom tertentu. Representasi diagram dari kariotipe suatu spesies disebut idiogram. Secara umum, dalam idiogram, kromosom dari kumpulan haploid suatu organisme diurutkan dalam urutan yang ukurannya semakin kecil.
Penggunaan kariotipe:
- Kariotipe dari kelompok yang berbeda kadang-kadang dibandingkan dan kesamaan dalam kariotipe dianggap menunjukkan hubungan evolusioner.
- Kariotipe juga menyarankan ciri primitif atau lanjutan dari suatu organisme. Kariotipe yang menunjukkan perbedaan besar antara kromosom terkecil dan terbesar dari kumpulan dan memiliki lebih sedikit kromosom metasentrik, disebut kariotipe asimetris, yang dianggap sebagai fitur yang relatif maju jika dibandingkan dengan kariotipe simetris yang memiliki semua kromosom metasentrik dengan ukuran yang sama. Levitzky (1931) mengemukakan bahwa pada tanaman berbunga terdapat kecenderungan yang menonjol ke arah kariotipe asimetris.
Bahan Kromosom:
Bahan kromosom adalah kromtin. Bergantung pada sifat pewarnaannya dengan pewarna dasar (khususnya reagen Feulgen), dua jenis kromatin berikut dapat dibedakan dalam inti interfase.
- Eukromatin:
Bagian kromosom yang ternoda ringan hanya terkondensasi sebagian; kromatin ini disebut eukromatin. Ini mewakili sebagian besar kromatin yang menyebar setelah mitosis selesai. Euchromatin mengandung gen struktural yang bereplikasi dan mentranskripsi selama fase G 1 dan S interfase. Ini dianggap kromatin aktif secara genetik, karena memiliki peran dalam ekspresi fenotipe gen. Dalam euchromatin, DNA ditemukan dikemas dalam serat 3 sampai 8 nm.
- Heterokromatin:
Di daerah pewarnaan gelap, kromatin tetap dalam keadaan kental dan disebut heterokromatin. Pada tahun 1928, Heitz mendefinisikannya sebagai daerah kromosom yang tetap terkondensasi selama interfase dan profase awal dan membentuk apa yang disebut chromocentre.
Heterokromatin dicirikan oleh kandungan sekuens DNA berulang yang sangat tinggi dan mengandung sangat sedikit, jika ada, gen struktural. Itu adalah replikasi yang terlambat (yaitu, itu direplikasi ketika sebagian besar DNA telah direplikasi) dan tidak ditranskripsi. Diperkirakan bahwa dalam heterokromatin, DNA dikemas rapat dalam serat 30 nm. Sekarang ditetapkan bahwa gen di wilayah heterokromatik tidak aktif.
Selama tahap profase awal dan pertengahan, daerah heterokromatik dibentuk menjadi tiga struktur yaitu kromomer, sentromer, dan kenop. Kromomer mungkin tidak mewakili heterokromatin sejati karena ditranskripsi.
Daerah sentromerik selalu mengandung timah heterokroma; di kelenjar ludah, daerah dari semua kromosom ini bergabung membentuk massa heterokromatik besar yang disebut kromosentre. Kenop adalah badan heterokromatin bulat, biasanya beberapa kali diameter kromosom yang bersangkutan, hadir dalam kromosom tertentu dari beberapa spesies, misalnya,
Jagung; kenop lebih jelas terlihat selama tahap pachytene pada jagung. Jika ada, kenop berfungsi sebagai penanda kromosom yang berharga.
Heterokromatin diklasifikasikan menjadi dua kelompok: (i) Konstitutif dan (itu) Fakultatif.
(i) Heterokromatin konstitutif tetap secara permanen dalam keadaan heterokromatik, yaitu tidak kembali ke keadaan eukromatik, misalnya daerah sentromerik. Ini berisi urutan pendek berulang DNA, yang disebut DNA satelit.
(ii) Heterokromatin fakultatif pada dasarnya adalah eukromatin yang telah mengalami heterokromatinisasi yang mungkin melibatkan satu segmen kromosom, seluruh kromosom (misalnya satu kromosom X manusia betina dan betina mamalia lain), atau satu set kromosom haploid utuh (misalnya, pada beberapa serangga, seperti kutu putih).
Komposisi kimia:
Kromatin terdiri dari DNA, RNA dan protein. Protein kromatin terdiri dari dua jenis: histon dan non-histon. Kromatin murni yang diisolasi dari inti interfase terdiri dari sekitar 30-40% DNA, 50-65% protein, dan 0,5-10% RNA: tetapi ada variasi yang cukup besar karena spesies dan jaringan dari spesies yang sama.
DNA:
Jumlah DNA yang ada dalam sel somatik normal suatu spesies adalah konstan untuk spesies itu; variasi apa pun dalam DNA dari nilai ini sangat berkorelasi dengan variasi yang sesuai pada tingkat kromosom. Gamet suatu spesies hanya mengandung setengah dari jumlah DNA yang ada dalam sel somatiknya. Jumlah DNA yang ada dalam sel somatik juga tergantung pada fase siklus sel.
Protein:
Protein yang terkait dengan kromosom dapat diklasifikasikan menjadi dua kelompok besar: (/) protein dasar atau histon dan (ii) protein non-histon.
Histon merupakan sekitar 80% dari total protein kromosom; mereka hadir dalam rasio hampir 1:1 dengan DNA (berat/berat). Berat molekulnya berkisar antara 10.000-30.000 dan sama sekali tidak mengandung triptofan. Histon adalah kelas protein yang sangat heterogen yang dapat dipisahkan dalam 5 fraksi berbeda yang ditetapkan sebagai H 1 H 2 a, H 2 b, H 3 dan H 4 setelah ewin (1975).
Fraksi H 1 kaya lisin, H 2 a dan H 2 b sedikit kaya lisin, sedangkan H 3 dan H 4 kaya arginin. Kelima fraksi ini ada di semua jenis sel eukariota, kecuali dalam sperma beberapa spesies hewan di mana mereka digantikan oleh kelas lain dari protein dasar molekul yang lebih kecil yang disebut protamine.
Histon memainkan fungsi utama dalam organisasi kromosom di mana H 2 a, H 2 b, H 3 dan H 4 terlibat dalam organisasi struktural serat kromatin, sedangkan fraksi H 1 menyatukan serat kromatin terlipat dari kromosom.
Protein non-histone membentuk sekitar 20% dari total massa kromosom, tetapi jumlahnya bervariasi dan tidak ada rasio pasti antara jumlah DNA dan non-histone yang ada dalam kromosom.
Mungkin ada 12 sampai lebih dari 20 jenis protein non-histon berbeda yang menunjukkan variasi dari satu spesies ke spesies lain dan bahkan dalam jaringan yang berbeda dari organisme yang sama. Kelas protein ini mencakup banyak enzim penting, seperti DNA dan RNA polimerase, dll.
Ultrastruktur Kromosom:
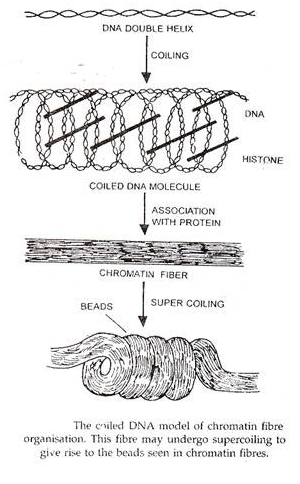
Studi mikroskop elektron telah menunjukkan bahwa kromosom memiliki fibril yang sangat halus dengan ketebalan 2 nm-4 nm. Karena lebar DNA adalah 2 nm, ada kemungkinan bahwa satu fibril berhubungan dengan satu molekul DNA. Beberapa model struktur kromosom telah diusulkan dari waktu ke waktu berdasarkan berbagai jenis data kromosom.
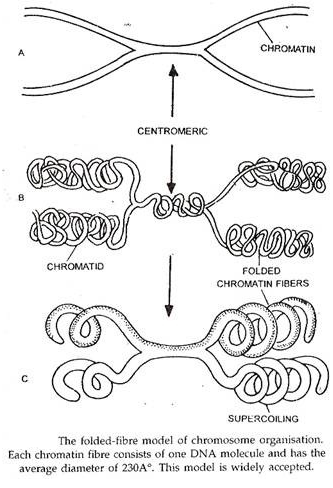
Model Serat Lipat dari Kromosom:
Model ini diusulkan oleh Du Praw pada tahun 1965 dan diterima secara luas. Menurut model ini, kromosom terdiri dari serat kromatin dengan diameter sekitar 230A°. Setiap serat kromatin hanya mengandung satu heliks ganda DNA yang dalam keadaan melingkar; kumparan DNA ini dilapisi dengan protein histon dan non-histon.
Dengan demikian serat kromatin 230A° diproduksi dengan menggulung heliks ganda DNA tunggal, yang gulungannya distabilkan oleh protein dan kation divalen (Ca ++ dan Mg ++ ). Setiap kromatid mengandung satu serat kromatin panjang; DNA dari serat ini bereplikasi selama interfase menghasilkan dua serat kromatin saudara, tetap tidak direplikasi di wilayah sentromerik sehingga kedua serat saudara tetap bergabung di wilayah tersebut.
Selanjutnya, serat kromatin juga mengalami replikasi di daerah sentromerik sehingga serat kromatin saudaranya terpisah di daerah ini juga. Selama pembelahan sel, dua serat kromatin mengalami pelipatan yang luas secara terpisah dengan cara yang tidak teratur untuk menghasilkan dua kromatid saudara.
Lipatan serat kromatin secara drastis mengurangi panjangnya dan meningkatkan kemampuan noda dan ketebalannya. Struktur terlipat ini biasanya mengalami supercoiling yang selanjutnya meningkatkan ketebalan kromosom dan mengurangi panjangnya. Sebagian besar bukti yang tersedia mendukung model ini.
Bukti luar biasa dari berbagai penelitian mendukung teori bahwa setiap kromatid mengandung satu molekul DNA raksasa. Bukti terkuat yang mendukung model unineme (kromatid beruntai tunggal) disediakan oleh studi tentang kromosom sikat lampu.
Organisasi Serat Kromatin:
Setiap model struktur serat kromatin harus memperhitungkan (i) pengemasan molekul DNA yang sangat panjang menjadi satu satuan panjang serat; (ii) produksi serat yang sangat tebal (230-300A 0 ) dari molekul DNA yang sangat tipis (20A°) dan (iii) ultrastruktur bead-on-a-string dari serat kromatin yang diamati khususnya selama replikasi. Dua model struktur serat kromatin yang jelas berbeda telah diusulkan:
- Model DNA Melingkar:
Ini adalah model organisasi serat kromatin yang paling sederhana dan diberikan oleh Du Praw. Menurut model ini, molekul DNA tunggal dari serat kromatin digulung dengan cara yang mirip dengan kawat pada pegas; gulungan yang disatukan oleh jembatan histon yang dihasilkan dengan mengikat molekul histon dalam alur besar molekul DNA. Struktur melingkar seperti itu yang akan distabilkan sebagai satu molekul histon akan mengikat beberapa gulungan DNA.
Struktur melingkar ini dilapisi dengan protein kromosom untuk menghasilkan struktur dasar serat kromatin (serat tipe A) yang dapat mengalami supercoiling untuk menghasilkan serat tipe B dari DuPraw yang mirip dengan manik-manik yang terlihat pada mikrograf elektron dari serat kromatin.
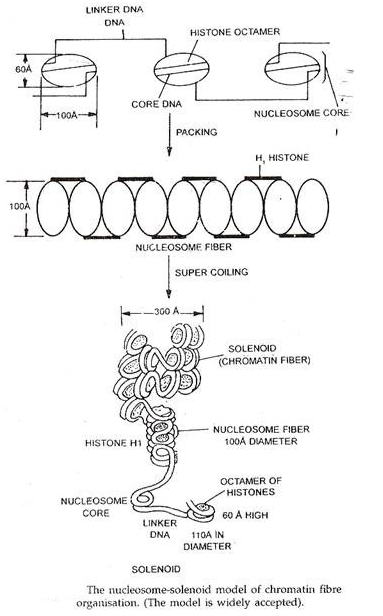
- Model Nukleosom-Solenoid:
Model ini dikemukakan oleh Romberg dan Thomas (1974) dan merupakan yang paling banyak diterima. Menurut model ini, kroma timah terdiri dari unit berulang yang disebut nukleosom. Nukleosom adalah partikel unit pengemas dasar dari kromatin dan memberikan kromatin penampilan “manik-manik-pada-tali” dalam mikrograf elektron yang membuka pengemasan tingkat tinggi (Olins dan Olins, 1974). Satu nukleosom lengkap terdiri dari inti nukleosom, DNA penghubung, rata-rata satu molekul histon H 1 dan protein kromosom somal terkait lainnya.
Inti Nukleosom:
Ini terdiri dari oktamer batu tinggi yang terdiri dari dua molekul, masing-masing histon H 2 a, H 2 b, H 3 dan H 4 . Selain itu, molekul DNA sepanjang 146 bp dililitkan di sekitar histone octamer ini dalam 13/4 putaran; segmen DNA ini tahan nuklease.
DNA penghubung:
Ukurannya bervariasi dari 8bp hingga 114 bp tergantung pada spesiesnya. DNA ini membentuk bagian benang dari serat kromatin manik-manik, dan rentan terhadap nuklease; dan manik-manik itu karena inti nukleosom. Jadi, DNA penghubung bergabung dengan dua nukleosom tetangga.
H 1 Histon:
Setiap nukleosom rata-rata mengandung satu molekul histon HI, meskipun distribusinya yang seragam di sepanjang serat kromatin tidak diketahui dengan jelas. Beberapa penelitian menunjukkan bahwa molekul histon H1 terlibat dalam menstabilkan superkoil serat kromatin nukleosom. Studi lain menunjukkan bahwa HI dikaitkan di bagian luar setiap inti nukleosom, dan bahwa satu molekul H1 menstabilkan sekitar 166 bp molekul DNA panjang.
Protein Kromosom Lainnya:
Baik DNA penghubung dan nukleosom dikaitkan dengan protein kromosom lainnya. Dalam kromatin asli, manik-manik berdiameter sekitar 110A°, tinggi 60A° dan berbentuk ellipsoidal. Setiap manik sesuai dengan inti nukleosom tunggal. Dalam beberapa kondisi, nukleosom berkumpul bersama tanpa DNA penghubung, yang menghasilkan serat kromatin setebal 100A° yang disebut serat nukleosom yang kemudian dapat menjadi superkoil untuk menimbulkan serat kromatin 300A° yang disebut solenoid.
Model nukleosom struktur serat kromatin konsisten dengan hampir semua bukti yang dikumpulkan sejauh ini.
Kromosom Khusus:
Beberapa jaringan organisme tertentu mengandung kromosom yang sangat berbeda dari yang normal dalam hal morfologi atau fungsi; kromosom tersebut disebut sebagai kromosom khusus. Jenis kromosom berikut dapat dimasukkan dalam kategori ini: (1) Kromosom sikat lampu, (2) Kromosom raksasa atau kromosom kelenjar ludah dan (3) Aksesori atau kromosom B.
Kromosom Lampbrush:
Kromosom lampbrush ditemukan di oosit dari banyak brata invertebrata dan semua vertebrata, kecuali mamalia; mereka juga telah dilaporkan pada ooosit manusia dan hewan pengerat. Tapi mereka telah dipelajari paling ekstensif dalam oosit amfibi.
Kromosom-kromosom ini paling jelas diamati selama tahap diploten yang berkepanjangan dari oo cytes . Selama diploten, kromosom homolog mulai terpisah satu sama lain, tetap berhubungan hanya pada beberapa titik sepanjang panjangnya.
Setiap kromosom dari suatu pasangan memiliki beberapa kromomer yang terdistribusi sepanjang panjangnya; dari masing-masing mayoritas kromomer umumnya sepasang loop lateral memanjang ke arah yang berlawanan tegak lurus terhadap sumbu utama kromosom.
Dalam beberapa kasus, lebih dari satu pasang, bahkan hingga 9 pasang loop dapat muncul dari satu kromomer. Lengkungan lateral ini membuat kromosom tampak seperti sikat lampu yang menjadi alasan untuk diberi nama ‘kromosom sikat lampu’.
Kromosom ini sangat panjang, dalam beberapa kasus panjangnya 800-1000^. Ukuran loop dapat berkisar dari rata-rata 9,5ja pada katak hingga 200|i pada newt. Pasangan loop diproduksi karena uncoiling dari dua serat kromatin (maka dua kromatid saudara) hadir dalam keadaan sangat melingkar dalam kromosom; ini membuat DNA mereka tersedia untuk transkripsi (sintesis RNA).
Jadi setiap loop mewakili satu kromatid kromosom dan terdiri dari satu heliks ganda DNA. Salah satu ujung setiap loop lebih tipis (thin end) dari ujung lainnya (thick end). Ada sintesis RNA yang luas di ujung tipis loop, sementara ada sedikit atau tidak ada sintesis RNA di ujung yang tebal.
Serat kromatin dari kromomer secara progresif diurai menuju ujung tipis dari sebuah gelung; DNA di wilayah ini mendukung sintesis RNA aktif tetapi kemudian dikaitkan dengan RNA dan protein menjadi lebih tebal.
DNA pada ujung yang tebal dari loop secara progresif ditarik dan dipasang kembali ke dalam kromomer. Jumlah pasang loop secara bertahap meningkat pada meiosis hingga mencapai maksimum pada diploten. Saat meiosis berlanjut, jumlah loop secara bertahap berkurang dan loop akhirnya menghilang karena disintegrasi daripada reabsorpsi kembali ke kromomer.
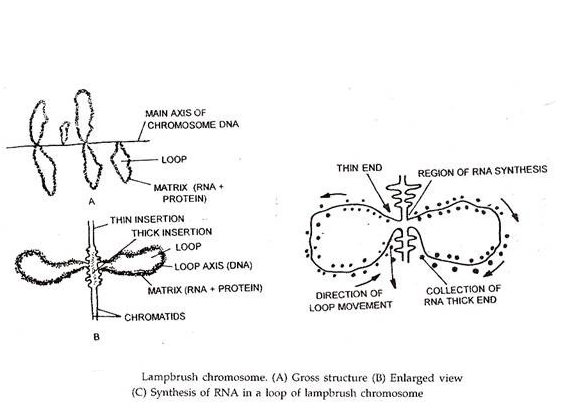
Loop mewakili situs aksi gen (transkripsi), dan fungsi kromosom sikat lampu adalah untuk menghasilkan sejumlah besar protein dan RNA yang disimpan dalam telur.
Kromosom Raksasa:
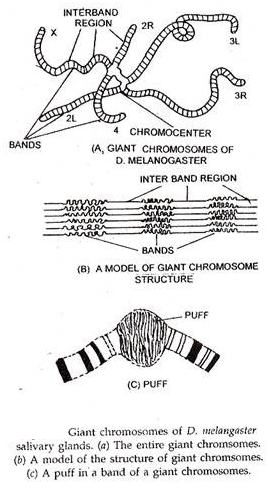
Kromosom raksasa ditemukan di jaringan tertentu, misalnya kelenjar ludah larva, epitel usus, tubulus Malphigian dan beberapa Diptera, misalnya Drosophila, Chironomous, Sciara, Rhyncosciara dll. Kromosom ini sangat panjang (hingga 200 kali ukurannya selama metafase mitosis di kasus Drosophila) dan sangat tebal, karenanya dikenal sebagai kromosom raksasa.
Mereka pertama kali ditemukan oleh Balbiani (1881) di kelenjar ludah dipteran, memberi mereka nama yang umum digunakan kromosom kelenjar ludah. Kromosom raksasa dipasangkan secara somatik. Akibatnya, jumlah kromosom raksasa ini di sel kelenjar air liur selalu tampak setengah dari sel somatik normal.
Kromosom raksasa memiliki pola pita transversal yang berbeda yang terdiri dari daerah kromatik dan akromatik alternatif. Pita kadang-kadang membentuk tiupan reversibel, yang dikenal sebagai tiupan kromosom atau cincin Balabiani, yang berhubungan dengan sintesis RNA aktif.
Kromosom raksasa mewakili seikat fibril yang muncul melalui siklus endoreduplikasi berulang (replikasi kromatin tanpa pembelahan sel) dari kromatid tunggal. Inilah mengapa kromosom ini juga dikenal sebagai kromosom polytene dan kondisinya disebut polyteny. Jumlah chromonemata (fibril) per kromosom dapat mencapai hingga 2000 dalam kasus ekstrim beberapa pekerja menempatkan angka ini setinggi 16.000.
Dalam D melanogaster, kromosom raksasa memancar sebagai lima lengan panjang dan satu lengan pendek dari massa amorf tunggal yang dikenal sebagai chromocentre. Chromocentre dibentuk oleh fusi daerah sentromerik dari semua kromosom dan, pada laki-laki, dari seluruh kromosom Y.
Lengan pendek yang memancar dari chromocentre mewakili kromosom IV, salah satu lengan panjang karena kromosom X, sedangkan empat lengan panjang lainnya mewakili lengan kromosom II dan III. Panjang total kromosom raksasa D. melanogaster sekitar 2000µ.
Kromosom tambahan:
Pada banyak spesies, ditemukan terlalu banyak kromosom tambahan selain komplemen somatik normal; kromosom ekstra ini disebut kromosom aksesori, kromosom B atau kromosom supernumerary.
Sekitar 600 spesies tumbuhan dan lebih dari 100 spesies hewan dilaporkan memiliki kromosom B. Kromosom B umumnya berukuran lebih kecil daripada kromosom komplemen somatik normal tetapi pada beberapa spesies ukurannya mungkin lebih besar (misalnya pada Sciara).
Salah satu fitur terpenting dari kromosom ini adalah bahwa jumlahnya dapat sangat bervariasi di antara individu dari spesies yang sama; pada jagung sebanyak 25-30 kromosom B dapat terakumulasi pada beberapa individu tanpa efek nyata pada fenotipnya. Kromosom ini umumnya diperoleh dan hilang dari individu suatu spesies tanpa efek merugikan atau menguntungkan yang nyata.
Namun, kehadiran beberapa kromosom B sering menyebabkan penurunan kekuatan dan kesuburan jagung. Dalam banyak kasus, sebagian besar bersifat heterokromatik, sementara pada beberapa spesies (misalnya jagung) sebagian bersifat heterokromatik, dan pada beberapa spesies lain (misalnya, Tradescantia) seluruhnya bersifat eukromatik. Mereka diyakini umumnya tidak aktif secara genetik, tetapi mereka mungkin tidak sepenuhnya tanpa gen.
Asal usul kromosom B pada sebagian besar spesies tidak diketahui. Pada beberapa hewan, mereka mungkin muncul karena fragmentasi kromosom Y heterokromatik. Pada jagung, ciri-ciri morfologis dan perilaku berpasangan kromosom B jelas menunjukkan bahwa mereka tidak memiliki segmen yang homolog dengan segmen kromosom komplemen somatik normal.
Kromosom B relatif tidak stabil; pada banyak spesies mereka cenderung dihilangkan dari jaringan somatik karena lagging dan non-disjunction dan mereka sering berubah morfologi melalui fragmentasi. Selanjutnya, mereka mungkin juga menunjukkan distribusi yang tidak teratur selama meiosis, tetapi mereka selalu dipertahankan dalam jaringan reproduksi.
Fungsi Kromosom:
Peran kromosom dalam hereditas dikemukakan secara terpisah oleh Sutton dan Bover pada tahun 1902. Ini dan berbagai fungsi kromosom lainnya dapat diringkas sebagai berikut.
- Diterima secara universal bahwa DNA adalah materi genetik, dan bahwa pada eukariota hampir semua DNA terdapat dalam kromosom. Dengan demikian, fungsi kromosom yang paling penting adalah menyediakan informasi genetik untuk berbagai fungsi seluler yang penting untuk pertumbuhan, kelangsungan hidup, perkembangan, reproduksi, dll., organisme.
- Fungsi lain yang sangat penting dari kromosom adalah melindungi materi genetik (DNA) agar tidak rusak selama pembelahan sel. Kromosom dilapisi dengan histon dan protein lain yang melindunginya dari kekuatan kimia (misalnya enzim) dan fisik.
- Sifat-sifat kromosom memastikan distribusi DNA (materi genetik) yang tepat ke inti anak selama pembelahan sel. Sentromer kromosom melakukan fungsi penting dalam pergerakan kromosom selama pembelahan sel yang disebabkan oleh kontraksi serat gelendong yang menempel pada daerah sentromerik kromosom.
- Tindakan gen pada eukariota diyakini diatur melalui protein histon dan non-histon yang terkait dengan kromosom.



