Mekanisme Penentuan Jenis Kelamin Di Bawah Kendali Genetik!
Penentuan jenis kelamin pada sebagian besar tumbuhan dan hewan berkaitan dengan studi tentang faktor-faktor yang bertanggung jawab untuk membuat individu jantan, betina, atau hermafrodit. Di masa lalu, mekanisme penentuan jenis kelamin dijelaskan murni berdasarkan kromosom seks, yang susunannya umumnya berbeda pada individu laki-laki dan perempuan.
Namun, dalam beberapa tahun terakhir, perbedaan telah dibuat pertama, antara penentuan jenis kelamin dan diferensiasi jenis kelamin dan kedua antara peran yang dimainkan oleh konstitusi kromosom dan gen spesifik (terletak pada kromosom seks dan autosom) dalam mencapai dimorfisme seksual.
Telah ditunjukkan bahwa penentuan jenis kelamin hanyalah sinyal yang memulai pola perkembangan laki-laki atau perempuan, dan bahwa diferensiasi jenis kelamin melibatkan jalur aktual peristiwa yang mengarah pada perkembangan tidak hanya organ laki-laki dan perempuan tetapi juga karakter seks sekunder.
- Mekanisme Penentuan Jenis Kelamin Dikendalikan Secara Genetik:
Mekanisme penentuan jenis kelamin di bawah kendali genetik pada dasarnya serupa pada tumbuhan dan hewan; berbagai mekanisme dapat diklasifikasikan ke dalam kategori berikut:
- Mekanisme kromosom seks atau heterogamesis
- Mekanisme keseimbangan jin
- Mekanisme haploidi atau hoplodiploidi pria
- Efek gen tunggal.
Mekanisme Kromosom Seks (Heterogamesis):
Pada sebagian besar hewan, individu jantan dan betina biasanya berbeda satu sama lain dalam hal jumlah atau morfologi homolog dari satu pasangan kromosom, pasangan ini disebut sebagai kromosom seks atau allosome.
Sebaliknya, kromosom yang jumlah dan morfologinya tidak berbeda antara jantan dan betina suatu spesies disebut autosom. Kromosom seks bertanggung jawab untuk penentuan jenis kelamin sedangkan autosom tidak memiliki hubungan dengan jenis kelamin dan mengandung gen yang menentukan karakter somatik individu.
Ada dua jenis kromosom seks: X dan Y. Kromosom X ditemukan pada pria dan wanita, meskipun satu jenis kelamin hanya memiliki satu sedangkan jenis kelamin lainnya memiliki dua kromosom X. Sebaliknya, kromosom Y biasanya hanya terdapat pada salah satu dari dua jenis kelamin suatu spesies, misalnya tikus jantan, Drospohila, manusia dll., dan burung betina, reptilia, dll.
Kromosom X memiliki sejumlah besar eukromatin dan sejumlah kecil heterokromatin. Euchromatin memiliki sejumlah besar bahan DNA, karenanya, banyak informasi genetik. Kromosom Y mengandung sejumlah kecil eukromatin dan sejumlah besar heterokromatin, sehingga hanya memiliki sedikit informasi genetik.
Jenis Mekanisme Kromosom Seks Penentuan Jenis Kelamin:
Dalam organisme diploidi dioecious mengikuti dua sistem penentuan kromosom seks seks telah diakui;
(a) Laki-laki heterogami
(b) Betina heterogami
Laki-laki Heterogami:
Pada jenis penentuan jenis kelamin kromosom seks, jenis kelamin perempuan memiliki dua kromosom X, sedangkan jenis kelamin laki-laki hanya memiliki satu kromosom X. Karena, laki-laki tidak memiliki kromosom X, oleh karena itu, selama gametogenesis menghasilkan dua jenis gamet, 50 persen gamet membawa mosomes kromosom X, sedangkan sisanya kekurangan kromosom X.
Seperti jenis kelamin yang menghasilkan dua jenis gamet yang berbeda dalam hal kromosom seks disebut seks heterogami. Oleh karena itu, jenis kelamin perempuan disebut jenis kelamin homogami. Laki-laki heterogami mungkin dari dua jenis berikut.
(i) Jenis XX-XO:
Pada serangga khususnya ordo Hemiptera (serangga sejati) dan Orthoptera (belalang dan kecoak) dan tumbuhan tertentu (misalnya, Vallisneria spiralis, Dioscorea sinuata, dll.), betina memiliki dua kromosom X (karenanya disebut XX) dan dengan demikian homogami, sedangkan laki-laki hanya memiliki satu kromosom X (maka disebut sebagai XO).
Kehadiran kromosom X yang tidak berpasangan menentukan jenis kelamin laki-laki. Laki-laki yang kekurangan satu kromosom X menghasilkan dua jenis sperma: setengah dari sperma memiliki satu kromosom X; sementara separuh lainnya tidak memilikinya. Penyatuan sperma yang memiliki kromosom X dengan sel telur menghasilkan zigot yang memiliki dua kromosom X (XX); zigot tersebut berkembang menjadi individu perempuan. Tetapi ketika sperma tanpa kromosom X membuahi sel telur, diperoleh zigot XO yang berkembang menjadi laki-laki. Jadi, setengah dari keturunan dari setiap perkawinan adalah perempuan, sedangkan separuh lainnya adalah laki-laki.
(ii) Tipe XX-XY:
Pada manusia, tikus, sebagian besar mamalia lainnya, Hemiptera, Coleoptera, Diptera (misalnya, Drosophila, lalat rumah dll.) beberapa ikan, beberapa amfibi dan pada tanaman angiospermia tertentu seperti Coccinia indica, album Melandrium, betina memiliki dua kromosom X homomorfik di sel tubuh mereka ((karenanya, disebut sebagai XX) dan mereka homogami, menghasilkan satu jenis telur, masing-masing dengan satu kromosom X.
Laki-laki dari organisme ini memiliki satu kromosom X dan satu kromosom Y (XY). Laki-laki yang memiliki dua kromosom seks heteromorfik menghasilkan dua jenis sperma: setengah dengan kromosom X dan setengah lagi dengan kromosom Y.
Pembuahan sel telur oleh sperma yang memiliki kromosom X menghasilkan zigot XX, yang berkembang menjadi betina. Tetapi zigot yang dihasilkan oleh penyatuan sel telur dengan sperma yang memiliki kromosom Y menghasilkan zigot XY yang menghasilkan laki-laki.
Betina Heterogami:
Dalam penentuan kromosom jenis kelamin jenis kelamin ini, jenis kelamin laki-laki memiliki dua kromosom X homomorfik, oleh karena itu, bersifat homogami dan menghasilkan satu jenis gamet, masing-masing membawa satu kromosom X.
Jenis kelamin perempuan terdiri dari satu kromosom X atau satu kromosom X dan satu kromosom Y. Jenis kelamin perempuan dengan demikian heterogami dan menghasilkan dua jenis telur, setengah dengan kromosom X dan setengah tanpa kromosom X (dengan atau tanpa kromosom Y.)
Betina heterogami mungkin dari jenis berikut:
(i) Sistem XO-XX (XO betina, XX jantan):
Sistem penentuan jenis kelamin ini dikenal pada beberapa spesies serangga, misalnya Fumea. Dalam spesies seperti itu, betina adalah heterogami (menghasilkan dua jenis telur, setengah dengan kromosom X dan setengah lagi tanpa kromosom X) dan laki-laki adalah jenis kelamin heterogami (memproduksi satu jenis sperma, yang masing-masing membawa satu kromosom X.
Penyatuan sperma dengan kromosom X yang mengandung sel telur menghasilkan zigot XX yang berkembang menjadi jantan. Tetapi pembuahan telur tanpa kromosom X dengan sperma menimbulkan zigot XO yang berkembang menjadi betina.
(ii) Sistem XY-XX (XY perempuan, XX laki-laki):
Sistem penentuan jenis kelamin ini beroperasi pada burung, reptil, beberapa serangga, misalnya ulat sutera, dll. Di sini betina memiliki konstitusi kromosom XY oleh karena itu merupakan jenis kelamin heterogami karena separuh telur memiliki X, sedangkan sisanya memiliki kromosom Y.
Laki-laki dari spesies ini memiliki dua kromosom X (XX); akibatnya, laki-laki adalah jenis kelamin homogami karena semua sperma yang diproduksi oleh laki-laki, memiliki satu kromosom X. Pembuahan sel telur yang mengandung X dengan sperma menimbulkan zigot XX, yang berkembang menjadi jantan. Zigot XY diproduksi ketika sel telur yang mengandung Y dibuahi oleh sperma, zigot tersebut berkembang menjadi betina.
- Penentuan jenis kelamin oleh Genie Balance:
Pada Drosophila, mencit, pria, dll., keberadaan kromosom XX dan XY biasanya diasosiasikan dengan keperempuanan dan kelelakian. Ketika pernyataan umum dibuat tentang situasi seperti itu, tampak seolah-olah kromosom spesifik (X dan Y) sendiri yang menentukan jenis kelamin zigot.
Namun, studi tentang aksi gen membuat kita berharap bahwa beberapa gen spesifik yang terletak di kromosom ini harus terlibat dalam penentuan jenis kelamin. Sebuah kesimpulan untuk efek ini dicapai oleh Bridges (1921) yang akhirnya mengajukan teori keseimbangan jin yang diakui dengan baik tentang penentuan jenis kelamin dalam Drosophila.
Pada tahun 1916, Bridges menemukan betina XXY dan jantan XO di Drosophila sambil mempelajari pewarisan gen mata vermilion yang terletak di kromosom X. Ini jelas menunjukkan bahwa konstitusi kromosom XX dan XY masing-masing tidak esensial untuk kewanitaan dan kelelakian, dan bahwa kromosom Y tidak berperan dalam penentuan jenis kelamin.
Beberapa saat kemudian, Bridges mendapatkan betina triploid; betina ini ketika dikawinkan dengan jantan diploid normal menghasilkan sejumlah situasi aneuploid. Dengan mengkorelasikan jenis kelamin individu dengan susunan kromosomnya, Bridges mengembangkan teori keseimbangan jin untuk penentuan jenis kelamin. Teori ini sepenuhnya menjelaskan mekanisme penentuan jenis kelamin pada Drosophila, dan kemungkinan besar juga berlaku untuk burung.
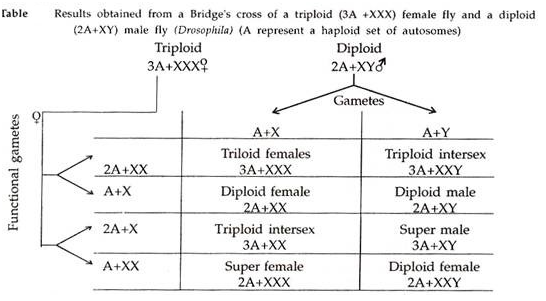
Teori keseimbangan genik menyatakan bahwa jenis kelamin seseorang ditentukan oleh keseimbangan antara gen kejantanan dan gen kewanitaan yang ada pada individu tersebut. Di Drosophila, gen untuk kejantanan ada di autosom, sedangkan untuk kewanitaan terletak di kromosom X.
Gen untuk kewanitaan yang ada dalam satu kromosom X lebih kuat daripada gen untuk kejantanan yang ada dalam satu set autosom, yaitu set autosom haploid. Kekuatan gen untuk kelelakian dan keperempuanan sangat seimbang sehingga ketika jumlah kromosom X dan kromosom autosom sama dalam satu individu, ia berkembang menjadi perempuan.
Seorang individu berkembang menjadi laki-laki hanya jika jumlah kromosom X-nya tepat setengah dari jumlah set autosomalnya. Intinya, jenis kelamin seseorang ditentukan oleh rasio jumlah kromosom X dan set autosomnya, rasio ini disebut sebagai indeks seks dan dinyatakan sebagai berikut:
Indeks seks = Jumlah kromosom X (X)/Jumlah set autosomal (A) = X/A
Seperti yang ditunjukkan pada tabel 5.2, ketika rasio X/A adalah 1,0, individu tersebut adalah perempuan dan jika rasionya adalah 0,50; itu akan laki-laki. Ketika keseimbangan ini terganggu, jenis kelamin individu menyimpang dari laki-laki normal atau perempuan normal. Misalnya, ketika rasio X/A turun antara 1,0 dan 0,50, itu akan menjadi interseks; ketika di bawah 0,50, itu akan menjadi laki-laki super dan ketika di atas 1,0, itu akan menjadi perempuan super.

Ginandromorf:
Konsep penentuan jenis kelamin yang dikembangkan untuk Drosophila diverifikasi oleh kemunculan gynandromorphs yang merupakan individu di mana bagian tubuh mengekspresikan karakter laki-laki, sedangkan bagian lain mengekspresikan karakter perempuan.
Fenotipe laki-laki pada ginandromor phs, pada satu ekstrem dapat meluas hingga sekitar separuh tubuh. Dalam beberapa lalat; bagian jantan dan betina berjalan membujur, sementara di beberapa bagian lainnya melintang. Gynandromorph selalu merupakan mosaik untuk kromosom X; bagian yang berfenotip jantan selalu XO, sedangkan yang berfenotipe betina XX.
Telah dikemukakan bahwa ginandromorf muncul dari zigot XX. Selama perkembangan embrio dalam satu sel atau lebih, salah satu dari dua kromosom tidak berpindah ke kutub mana pun pada anafase dan akibatnya hilang. Akibatnya, satu atau lebih sel anak yang memiliki satu kromosom X diproduksi; sel-sel ini membelah dan memunculkan bagian laki-laki ginandromorf.
Jika distribusi kromosom X yang tidak teratur terjadi pada pembelahan mitosis pertama pada zigot, fenotipe laki-laki akan meluas hingga tepat setengah dari tubuh ginandromorf. Oleh karena itu, luas bagian jantan dalam ginandromorf bergantung pada tahap perkembangan embrionik ketika distribusi kromosom X yang tidak teratur terjadi; semakin dini terjadinya, semakin besar ukuran porsi jantan.
Penentuan Jenis Kelamin pada Gen Y-linked:
Pada mamalia, axolotl (suatu Amphibia) dan beberapa tumbuhan, misalnya Melandrium, kromosom Y sangat penting untuk perkembangan kelelakian. Pada manusia XO, XX, XXX dan XXXX individu mengembangkan fenotipe perempuan. Tetapi dengan adanya satu kromosom Y, individu-individu ini, yaitu XY, XXY, XXXY dan XXXXY, mengembangkan fenotipe laki-laki.
Jadi satu kromosom Y cukup untuk mengatasi efek dari empat kromosom X dan menghasilkan fenotipe laki-laki. Tetapi manusia normal perempuan dan laki-laki masing-masing hanya diproduksi oleh konstitusi kromosom XX dan XY.
Kondisi XO menghasilkan sindrom Turner (wanita mandul), sedangkan XXY mengarah pada perkembangan sindrom Klinefelter (pria mandul). Kapasitas penentuan laki-laki tampaknya terletak di lengan pendek kromosom Y manusia (Y s ); penghapusan Y s memungkinkan pengembangan fenotipe wanita normal bahkan pada individu XY. Dalam kasus yang jarang terjadi, manusia XX dan tikus XX menunjukkan fenotip jantan.
Pada sebagian besar mamalia, ekspresi antigen, antigen HY, berkorelasi erat dengan perkembangan testis. Bahkan laki-laki XX menunjukkan antigen HY, sedangkan perempuan XY tidak memiliki antigen ini. Gen yang mengatur keberadaan antigen HY terletak di lengan pendek kromosom Y. Pada tikus, lengan pendek kromosom Y mengandung daerah sex reversal (sxr) yang penting untuk perkembangan kelelakian.
AKU AKU AKU. Mekanisme Haploidi Pria atau Haplodiploidi:
Partenogenesis haploidi atau haplodiploidi jantan atau arrhenotokous sangat umum terjadi pada serangga hymenopterous seperti semut, lebah, dan tawon (misalnya, Braco?i hebetor). Pada serangga ini, telur yang telah dibuahi berkembang menjadi betina diploid dan yang tidak dibuahi menjadi jantan haploid; jadi arrhenotoky adalah bentuk reproduksi dan alat penentuan jenis kelamin.
Selama spermatogenesis, semua n kromosom laki-laki secara teratur berpindah ke satu kutub pada anafase-I sehingga kutub yang berlawanan tidak menerima kromosom sama sekali. Jadi semua sperma biasanya haploid. Meiosis normal selama oogenesis menghasilkan semua telur haploid.
Pembuahan telur menghasilkan zigot diploid yang berkembang menjadi larva diploid. Biasanya, larva seperti itu melahirkan pekerja, yaitu betina mandul. Tetapi larva diploid yang diberi royal jelly (mengandung madu, serbuk sari dan beberapa zat yang dikeluarkan oleh pekerja) berkembang menjadi betina subur yang disebut ratu. Di sisi lain, telur yang tidak dibuahi berkembang secara partenogenik untuk menghasilkan larva haploid dan pada akhirnya jantan haploid yang sepenuhnya subur disebut drone.
- Kontrol Gen Tunggal Seks:
Pada banyak hewan, gen autosom tunggal mengesampingkan efek kromosom seks yang ada pada individu. Gen ini umumnya resesif, tetapi dalam beberapa kasus mungkin dominan.
Contoh klasik dari gen semacam itu adalah gen transfer resesif autosomal (tra) dari Drosophila. Ketika gen ini hadir dalam keadaan homozigot (tratra), ia mengubah zigot XX untuk berkembang menjadi jantan yang mandul. Gene tra tidak berpengaruh baik pada laki-laki atau ketika berada dalam keadaan heterozigot pada perempuan.
Ketika betina heterozigot untuk tra (XX Tra tra) dikawinkan dengan jantan homozigot untuk tra (XY tra tra), hanya 25% keturunannya yang betina, sedangkan sisanya 75% adalah jantan. Sepertiga dari laki-laki, bagaimanapun, adalah individu XX mandul yang homozigot untuk tra; mereka diubah menjadi kejantanan oleh gene tra.
Gen resesif serupa, mungkin autosomal, feminisasi testis menginduksi manusia XY untuk mengembangkan payudara dan vagina; individu seperti itu, bagaimanapun, memiliki testis yang merosot dan steril. Gen ini tidak mempengaruhi karakteristik individu wanita.



