Bagian yang berbeda dari sel bakteri umum telah ditunjukkan pada Gambar 2.3 dan telah dijelaskan sebagai berikut:
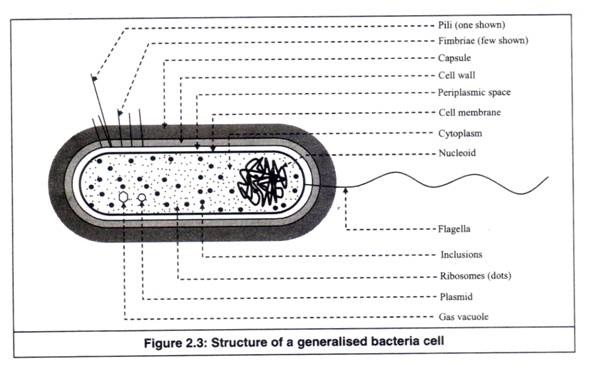
1. Flagela:
Flagela bakteri adalah pelengkap heliks seperti rambut berfilamen tipis yang menonjol melalui dinding sel dan bertanggung jawab atas motilitas bakteri. Sebagian besar bakteri motil memiliki flagela.
Panjangnya sekitar 10-15µ. Flagela bakteri sama sekali berbeda dari flagela eukariotik dalam struktur dan mekanisme kerjanya. Mereka jauh lebih tipis daripada flagela atau silia sel eukariotik. Tidak ada membran sel yang hadir di atas flagela bakteri tidak seperti pada flagela eukariotik.
Selain itu, flagela bakteri, tidak seperti flagela eukariotik, agak kaku dan tidak bergerak bolak-balik, melainkan berputar seperti baling-baling perahu untuk pergerakan sel. Flagela bakteri memiliki tiga bagian yaitu; (a) Basal Body, (b) Hook dan (c) Filament (Gambar 2.4).
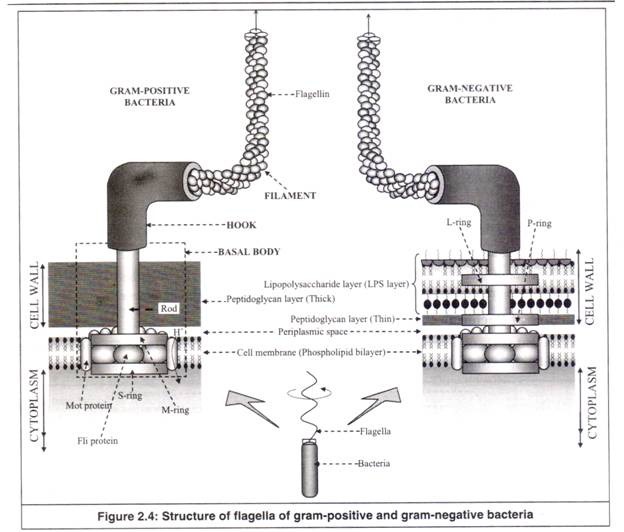
(sebuah) Tubuh Basal:
Ini membentuk bagian dasar flagel dan berlabuh di membran sitoplasma dan dinding sel. Ini memberikan jangkar ke flagela serta berfungsi sebagai motor, sehingga memberikan gerakan berputar dari flagel.
Ini terdiri dari batang pusat kecil yang melewati sistem cincin. Dalam kasus bakteri gram positif, di mana dinding sel terdiri dari lapisan peptidoglikan saja, tubuh basal hanya memiliki sepasang cincin; cincin-M dan cincin-S, keduanya tertanam di membran sel.
Di sekeliling cincin ini terdapat kelompok protein yang disebut protein mot. Pergerakan proton melintasi membran sel melalui protein mot ini menghasilkan gaya gerak proton (energi) yang diperlukan untuk perputaran kedua cincin ini, yang pada akhirnya bertindak sebagai motor untuk perputaran seperti baling-baling flagela.
Kelompok protein lain, yang disebut protein fli, hadir terjepit di antara dua cincin ini. Mereka berfungsi sebagai sakelar motor, membalikkan rotasi flagela sebagai respons terhadap sinyal intraseluler.
Dalam kasus bakteri gram negatif, yang dinding selnya terdiri dari lapisan luar lipopolisakarida (LPS) selain lapisan peptidoglikan dalam yang lebih tipis, badan basal memiliki sepasang cincin lagi; cincin-L tertanam di lapisan LPS dan cincin-P di lapisan peptidoglikan tipis.
(b) Kait:
Kait pendek menghubungkan tubuh basal ke filamen flagel. Ini sedikit lebih tebal dari filamen. Aksi motorik tubuh basal ditransmisikan ke filamen melalui pengait.
(c) Filamen:
Ini adalah bagian flagela seperti rambut panjang, yang muncul dari kail. Ini beberapa kali lebih lama dari sel bakteri. Itu terbuat dari satu jenis protein, ‘flagellin’ yang berat molekulnya sekitar 40.000. Molekul flagellin membentuk rantai; tiga rantai tersebut terjalin untuk membentuk flagel. Tidak seperti rambut yang tumbuh di pangkalnya, filamen flagel tumbuh di ujungnya.
Asam amino melewati pusat berongga filamen dan menambah ujung distalnya. Proses ini disebut self-assembly, karena semua informasi untuk struktur akhir filamen berada di dalam subunit protein itu sendiri. Filamennya agak kaku dan tidak bergerak bolak-balik, melainkan berputar seperti baling-baling perahu untuk pergerakan sel.
Spirochetes tidak memiliki flagela eksternal. Namun, mereka bergerak dengan bantuan struktur mirip flagela yang terletak di dalam sel mereka tepat di bawah selubung luar. Ini disebut flagela periplasma, flagela aksial, fibril aksial atau endoflagella.
Flagela disusun dengan cara yang berbeda pada bakteri yang berbeda. Susunan flagella pada sel bakteri disebut flagellation. Flagellation adalah dari empat jenis berikut (Gambar 2.5).
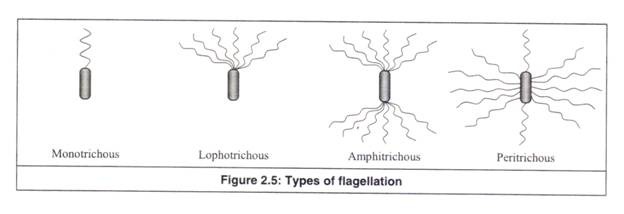
(a) Flagel tunggal (Mono: tunggal; trichous: rambut): Sebuah flagel tunggal hadir di salah satu ujung sel.
(b) Lophotrichous flagellation (Lopho: jumbai; trichous: rambut): Seberkas atau sekelompok flagela hadir di salah satu ujung sel.
(c) Pencambukan amfitrichous (Amphi: keduanya; trichous: rambut): Flagella ada di kedua ujung sel baik secara tunggal atau berjambul.
(d) Pencambukan peritrichous (Peri: sekitar; trichous: rambut): Flagela ada di seluruh permukaan sel.
2. Fimbria:
Fimbriae adalah pelengkap seperti rambut berfilamen tipis, yang secara struktural mirip dengan flagela, tetapi tidak terlibat dalam motilitas bakteri (Gambar 2.3). Tidak seperti flagela, mereka jauh lebih pendek dan lebih banyak, tetapi seperti flagela, mereka terbuat dari molekul protein.
Fimbriae hadir dalam beberapa spesies bakteri, yang membantu mereka menempel pada sel organisme lain dan mencegahnya hanyut oleh aliran lendir atau cairan tubuh. Dengan demikian, ini sangat membantu bakteri patogen untuk membuat infeksi di tubuh inang. Mereka juga membantu membentuk pelikel atau biofilm pada permukaan.
3.Pili:
Pili adalah pelengkap seperti rambut berfilamen tipis, yang secara struktural mirip dengan flagela, tetapi tidak terlibat dalam motilitas bakteri (Gambar 2.6). Mereka juga secara struktural mirip dengan fimbriae, tetapi umumnya lebih panjang dan hanya satu atau beberapa pili yang ada di permukaan sel bakteri.
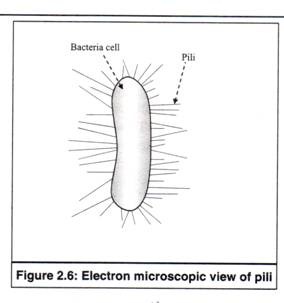
Itu terbuat dari satu jenis protein, ‘pilin’, yang berat molekulnya sekitar 17.000. Molekul pilin disusun secara heliks untuk membentuk satu filamen kaku dengan inti berongga di tengah. Pada beberapa bakteri, materi genetik ditransfer melalui inti berongga pili selama konjugasi (kawin dengan bakteri lain).
Pili semacam itu disebut pili kesuburan (F pili) atau pili seks. Pada beberapa bakteri patogen, pili bertindak mirip dengan fimbriae dalam perlekatan pada sel inang dan pembentukan infeksi. Aksi aglutinasi pili menghasilkan pembentukan film bakteri yang terlihat pada permukaan biakan kaldu.
4. Lapisan-S:
Lapisan-S (lapisan permukaan) adalah lapisan yang terdapat pada permukaan sebagian besar bakteri patogen, yang terdiri dari susunan protein dua dimensi dengan tampilan kristal dalam berbagai simetri, seperti heksagonal, tetragonal, atau trimerik (Gambar 2.7) .
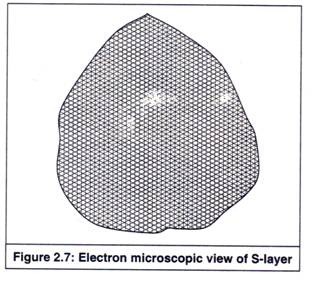
Simetri kristal bergantung pada komposisi protein lapisan-S. Ini terkait dengan struktur dinding sel (dengan lapisan peptidoglikan pada bakteri gram positif dan dengan lapisan LPS pada bakteri gram negatif).
Ini berfungsi sebagai penghalang permeabilitas eksternal, memungkinkan lewatnya zat dengan berat molekul rendah dan tidak termasuk zat dengan berat molekul tinggi. Ini juga memberikan perlindungan pada bakteri patogen, melawan mekanisme pertahanan inang.
5. Kapsul (lapisan lendir atau glikokaliks):
Banyak bakteri memiliki bahan kental berlendir atau bergetah di luar sel, mengelilingi dinding sel (Gambar 2.8). Itu terbuat dari makromolekul, yang tidak terikat secara kovalen dengan bakteri.
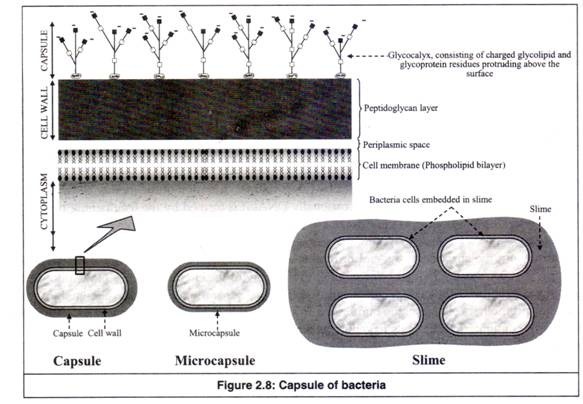
Makromolekul permukaan ini terutama bermuatan polisakarida (glikoprotein atau glikolipid) dan oleh karena itu disebut ‘glikokaliks’ (gliko: karbohidrat; kelopak: bagian bunga di bawah kelopak yang memanjang keluar dari tangkai). Glycocalyx didefinisikan sebagai bahan yang mengandung polisakarida yang terletak di luar sel sebagai jaringan longgar serat polimer yang memanjang keluar dari sel.
Mereka disintesis dan disekresikan oleh bakteri, yang berikatan dengan reseptor pada permukaan jaringan inang. Glycocalyx tidak hanya ditemukan di sekitar beberapa sel bakteri; itu juga ditemukan di sekitar beberapa sel tumbuhan dan hewan.
Glycocalyx dapat terdiri dari tiga jenis sebagai berikut:
(a) Kapsul:
Ini adalah lapisan polimer yang terdiri dari lapisan kental yang padat dan terdefinisi dengan baik yang mengelilingi sel bakteri di banyak spesies. Lapisan kental dapat divisualisasikan dengan mikroskop cahaya menggunakan teknik pewarnaan khusus.
Serat polimer glikokaliks diatur dalam matriks ketat yang tidak termasuk partikel seperti tinta India. Bakteri yang berkapsul disebut bakteri berkapsul, sedangkan yang tidak berkapsul disebut bakteri tidak berkapsul.
(b) Mikrokapsul:
Ini adalah lapisan kental yang mengelilingi sel bakteri, yang sangat tipis sehingga tidak dapat divisualisasikan dengan mikroskop cahaya.
(c) Lendir:
Ini adalah massa serat polimer yang menyebar yang tampaknya tidak terikat pada sel tunggal mana pun. Lapisan kental sangat tebal sehingga banyak sel yang tertanam dalam matriks lendir. Lapisan mudah berubah bentuk dan tidak mengecualikan partikel. Sulit untuk divisualisasikan dengan mikroskop cahaya.
Fungsi kapsul dan lendir diberikan di bawah ini:
- Mencegah pengeringan sel, dengan mengikat molekul air.
- Mencegah perlekatan bakteriofag (virus yang menyerang bakteri) pada sel bakteri.
- Ini memberikan virulensi (kemampuan menghasilkan penyakit) dengan menghambat pengenalan mereka dan penghancuran selanjutnya oleh fagosit (sel darah putih atau sel darah putih).
- Membantu bakteri patogen untuk perlekatannya pada reseptor di permukaan jaringan inang agar terinfeksi.
- Ini memberikan kepatuhan antara sel bakteri yang berbeda.
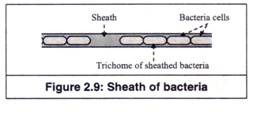
6. Selubung:
Selubung adalah struktur tubular berongga panjang, yang membungkus rantai sel bakteri membentuk ‘trikoma’ (Gambar 2.9). Bakteri berselubung umum ditemukan di habitat air tawar yang kaya akan bahan organik, seperti sungai yang tercemar, trickling filter, dan pencerna lumpur aktif di instalasi pengolahan limbah. Selubung dapat dengan mudah divisualisasikan, ketika beberapa sel, yang disebut ‘sel swarmer flagellated’ dari trikoma bergerak keluar dari selubung meninggalkan selubung kosong.
7. Kelebihan:
Pros-thecae adalah ekstensi sitoplasma dari sel bakteri, yang membentuk pelengkap seperti hifa, tangkai atau kuncup. Diameternya lebih kecil dari sel dewasa, mengandung sitoplasma dan dibatasi oleh dinding sel. Bakteri yang memiliki prosthecae disebut bakteri prosthecate.
Berbeda dengan pembelahan sel pada bakteri lain, yang terjadi melalui pembelahan biner yang menghasilkan dua sel bakteri anak yang setara, di prosthecates melibatkan pembentukan sel anak baru dengan sel induk mempertahankan identitasnya setelah pembelahan sel selesai.
‘Hifa’ adalah pelengkap seluler yang dibentuk oleh perpanjangan seluler langsung dari sel induk yang mengandung dinding sel, membran sel, ribosom, dan kadang-kadang DNA. Ini dibentuk oleh pemanjangan pertumbuhan kecil sel bakteri pada satu titik.
Pertumbuhan seperti itu pada satu titik disebut ‘pertumbuhan kutub’ berbeda dengan ‘pertumbuhan interkalar’ yang terjadi pada semua bakteri lain, di mana pertumbuhan bakteri terjadi di seluruh permukaan. Pada ujung hifa, terbentuk kuncup yang membesar dan menghasilkan flagel.
DNA dalam sel induk bereplikasi dan salinan DNA sirkular dipindahkan sepanjang hifa ke dalam kuncup. Sebuah septum silang kemudian terbentuk memisahkan tunas yang masih berkembang dari hifa dan sel induk.
Sekarang, kuncup pecah dari sel induk dan berenang menjauh sebagai sel anak. Kemudian sel anak kehilangan flagelnya dan secara bertahap tumbuh menjadi sel induk yang siap membentuk hifa dan kuncup.
‘Tangkai’ adalah pelengkap seluler yang digunakan oleh bakteri air tertentu untuk menempel pada substrat padat. Di ujung tangkai ada struktur yang disebut ‘pegangan’, dimana tangkai melekat pada substratum. Selain menyediakan perlekatan, tangkai juga meningkatkan luas permukaan sel bakteri, yang membantunya dalam penyerapan nutrisi dan pembuangan limbah.
Pada beberapa bakteri, tangkai beberapa individu tetap menempel membentuk ‘mawar’. Pembelahan sel pada bakteri bertangkai terjadi dengan pemanjangan sel dengan arah berlawanan dengan arah tangkai diikuti dengan pembelahan. Sebuah flagel tunggal terbentuk di tiang di seberang tangkai.
Sel anak berflagel yang terbentuk disebut ‘swarmer’. Ini terpisah dari sel induk yang tidak bertanda, berenang di sekitar dan menempel pada permukaan baru membentuk tangkai baru di kutub yang bertanda. Flagel kemudian hilang.
‘Tunas’ adalah pelengkap seluler, yang langsung berkembang sebagai sel anak dari sel induk tanpa campur tangan pembentukan hifa.
8. Dinding Sel:
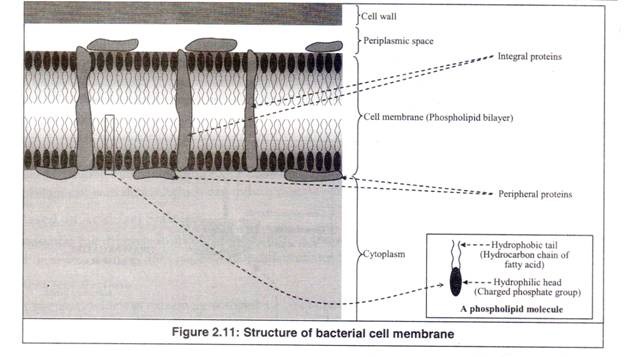
Dinding sel adalah struktur kaku yang langsung mengelilingi membran sel sel bakteri. Selaput sel bakteri adalah bilayer fosfolipid yang mirip dengan semua sel hewan dan tumbuhan lainnya (Gambar 2.11).
Sementara sel hewan tidak memiliki dinding sel, sel tumbuhan dan sel bakteri memiliki dinding sel yang mengelilingi membran sel. Namun, tidak seperti dinding sel sel tumbuhan yang sebagian besar tersusun atas selulosa, dinding sel sel bakteri sebagian besar tersusun atas lapisan peptidoglikan (Gambar 2.10).
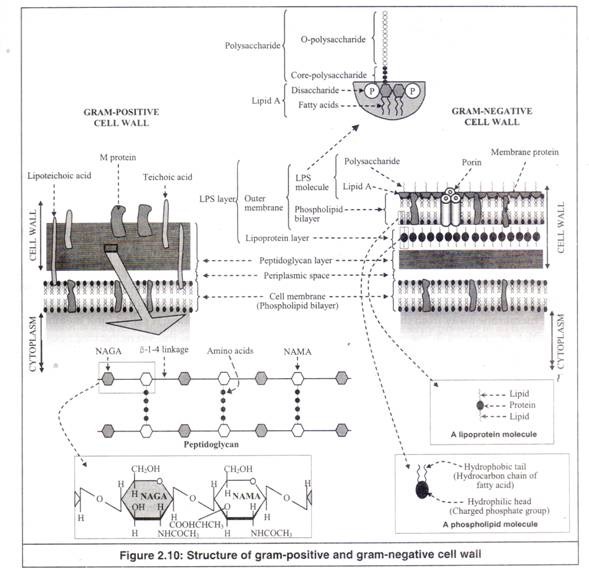
Sebelumnya, peptidoglikan juga disebut murein atau mucopeptide. Peptidoglikan terutama terdiri dari asam amino dan gula (peptido: rantai asam amino; glikan: gula). Dua gula yaitu N-acetylglucosamine (NAGA) dan N-acetylmuramic acid (NAMA) terikat satu sama lain secara bergantian melalui ikatan glikosidik (β-1-4 linkage) untuk membentuk rantai gula paralel yang panjang.
Terlampir pada molekul NAMA adalah rantai samping pendek dari empat atau lima asam amino yang terikat satu sama lain melalui ikatan peptida. Biasanya asam amino adalah alanin, asam glutamat dan asam lisin atau diaminopimelic (DAP). Peptida pendek ini membentuk ikatan silang antara rantai gula yang berdekatan untuk membentuk jaringan atau lapisan.
Meskipun ikatan glikosidik dalam rantai gula sangat kuat, ikatan ini saja tidak dapat memberikan kekakuan ke segala arah. Kekakuan tinggi diperoleh, hanya ketika rantai ini dihubungkan silang oleh ikatan peptida dalam peptida pendek penghubung silang.
Komposisi dinding sel tidak sama untuk semua bakteri; melainkan ada perbedaan yang luar biasa dalam komposisinya, berdasarkan pengelompokan bakteri seperti yang diberikan di bawah ini:
A. Berdasarkan Pewarnaan Gram:
Berdasarkan perbedaan komposisi dinding sel, teknik pewarnaan khusus yang disebut ‘pewarnaan gram’, dinamai Dr. Christian Gram yang mengembangkannya, membedakan bakteri menjadi dua kelompok sebagai berikut:
(1) Bakteri Gram-positif:
Setelah pewarnaan bakteri dengan kristal violet, jika dinding selnya menolak penghilangan warna dengan mencucinya dengan zat penghilang warna (etanol atau aseton), itu adalah bakteri gram positif. Contoh: Bacillus, Staphylococcus.
(2) Bakteri Gram-negatif:
Setelah pewarnaan bakteri dengan kristal violet, jika dinding selnya memungkinkan untuk mengalami penghilangan warna dengan mencucinya dengan zat penghilang warna (etanol atau aseton), itu adalah bakteri gram negatif. Contoh: Escherichia, Salmonella, Vibrio.
Diferensiasi bakteri menjadi kelompok gram positif dan gram negatif memberikan petunjuk paling penting untuk melangkah lebih jauh ke arah yang tepat untuk identifikasi bakteri yang tidak diketahui.
Dinding Sel Gram-Positif:
Dinding sel gram positif sangat sederhana. Ini terutama terbuat dari lapisan peptidoglikan yang tebal, yang merupakan sekitar 90% dari dinding sel. Lapisan tebal ini membuat bakteri gram positif tahan terhadap lisis osmotik.
Dinding juga mengandung asam teichoic, asam lipoteichoic dan protein M, yang merupakan antigen permukaan utama dari dinding sel gram positif. Asam teikoat bermuatan negatif dan sebagian bertanggung jawab atas muatan negatif permukaan sel bakteri. Ini juga membantu dalam perjalanan ion melalui dinding sel.
Asam lipoteikoat adalah asam teikoat, yang terikat pada lipid membran. Protein M membantu beberapa bakteri dalam menyebabkan penyakit dengan mencegah mereka ditelan oleh WBC. Antibiotik, penisilin (diproduksi oleh jamur, Penicillium notatum) dan sefalosporin membunuh bakteri gram positif dengan mencegah sintesis peptidoglikan. Lisozim (enzim yang ditemukan dalam air mata, air liur, albumin telur, dan lendir ikan) membunuh bakteri gram positif dengan memutus ikatan glikosidik (hubungan β-1-4) antara NAGA dan NAMA.
Dinding Sel Gram-Negatif:
Dinding sel gram negatif sangat kompleks dan lebih rapuh daripada dinding sel gram positif. Ini memiliki lapisan tipis peptidoglikan, yang hanya membentuk sekitar 10% dari dinding sel. Itu dikelilingi oleh lapisan pelindung tebal yang disebut lapisan lipopolisakarida (LPS), yang menahan penetrasi beberapa bahan kimia yang berpotensi beracun ke dalam sel bakteri.
Lapisan LPS terdiri dari 1) lapisan lipoprotein dan 2) membran luar. Satu permukaan lapisan lipoprotein langsung melekat pada lapisan peptidoglikan dan permukaan lainnya meluas ke membran luar.
Ini berfungsi sebagai jangkar antara lapisan peptidoglikan dan membran luar. Membran luar adalah bilayer fosfolipid tertanam protein dengan struktur ‘unit membran’ mirip dengan membran sel. Daun bagian dalam membran luar adalah lapisan molekul fosfolipid saja.
Namun, tidak seperti membran satuan, beberapa molekul fosfolipid di daun terluar digantikan oleh molekul LPS. Setiap molekul LPS terbuat dari bagian lipid yang disebut ‘lipid A’ yang secara kovalen terkait dengan polisakarida seperti rambut.
Ketika bakteri patogen gram negatif terbunuh di dalam tubuh inang, lipid A, yang merupakan komponen internal bakteri gram negatif, dilepaskan, menimbulkan reaksi toksik dan gejala penyakit. Itu sebabnya, lipid A disebut juga ‘endotoksin’ (endo: internal).
Sebaliknya, ‘eksotoksin’ adalah zat beracun yang disintesis di dalam sel bakteri dan dilepaskan ke luar, yang menyebabkan penyakit, jika ada di dalam atau masuk ke dalam tubuh inang. Tidak seperti lipid normal lainnya seperti trigliserida, yang terbuat dari tiga asam lemak yang terikat pada gliserol, lipid A terbuat dari asam lemak yang terikat pada disakarida yang terdiri dari dua NAGA fosfat.
Asam lemak dalam lipid A biasanya asam kaproat, laurat, miristat, palmitat, dan stearat. Polisakarida, yang terikat pada lipid A di ujung lain disakarida, terdiri dari dua bagian; 1) polisakarida inti dalam dan 2) polisakarida O luar.
Membran luar juga mengandung protein yang disebut ‘porin’, yang mengandung tiga subunit identik. Setiap subunit berbentuk silinder dengan pori pusat, tempat bahan dapat lewat. Dengan demikian, porin membuat membran luar relatif permeabel.
Porin terdiri dari dua jenis; 1) porin non-spesifik, yang membentuk saluran berisi air untuk dilewati zat kecil jenis apa pun dan 2) porin spesifik, yang memiliki tempat pengikatan khusus untuk dilewati zat tertentu.
Beberapa bahan (larutan, jarum suntik, jarum, dll.), meskipun dibuat bebas dari mikroba patogen dengan sterilisasi panas, masih mempertahankan sisa-sisa dinding sel bakteri gram negatif yang dihasilkan dengan membunuhnya. Jika masuk ke tubuh inang, puing-puing dinding sel ini dapat menyebabkan gejala penyakit. Puing-puing dinding sel yang rusak ini, yang dapat menyebabkan gejala penyakit, disebut ‘pirogen’.
B. Berdasarkan Pewarnaan Tahan Asam:
Berdasarkan perbedaan komposisi dinding sel, teknik pewarnaan khusus lainnya yang disebut ‘pewarnaan tahan asam’ membedakan bakteri menjadi dua kelompok sebagai berikut:
(1) Bakteri Tahan Asam:
Bakteri, yang sangat sulit untuk diwarnai, tetapi sekali diwarnai, sama sulitnya untuk menghilangkan noda dari selnya, bahkan dengan penggunaan asam-alkohol yang kuat sebagai agen penghilang warna, adalah bakteri tahan asam (asam-mencintai bakteri).
Contoh: Mycobacterium spp. [M. tuberculosis (bakteri TBC), M. leprae (bakteri kusta), M. smegmatis (bakteri alami smegma), M. marinum (bakteri TBC ikan laut)]. Mereka memiliki dinding sel lilin tebal yang terbuat dari bahan lipid.
(2) Bakteri tidak tahan asam:
Bakteri yang mudah diwarnai dan juga mudah didekolorisasi oleh asam-alkohol sebagai agen penghilang warna merupakan bakteri yang tidak tahan asam (non-acid-loving bacteria). Contoh: Semua bakteri lain kecuali Mycobacterium spp. Pada bakteri ini, dinding selnya tidak setebal lilin.
Diferensiasi bakteri menjadi kelompok tahan asam dan tidak tahan asam memberikan petunjuk paling penting untuk identifikasi Mycobacterium spp.
Varian yang kekurangan dinding:
Beberapa bakteri secara alami ada tanpa dinding sel. Mycoplasma, yang merupakan bakteri terkecil, tidak memiliki dinding sel. Ini dilindungi dari lisis osmotik dengan adanya sterol dalam membran sel dan dengan mengadopsi keberadaan parasit dalam lingkungan host eukariotik yang menguntungkan secara osmotik.
Demikian pula, beberapa bakteri laut kekurangan dinding sel, di mana lisis osmotik dicegah oleh konsentrasi garam air laut yang tinggi. Sebagian besar bakteri gram positif kehilangan dinding selnya (peptidoglikan) dengan adanya lisozim.
Namun, sel mereka bertahan hidup jika berada di lingkungan isotonik seperti luka berisi nanah. Bakteri yang kekurangan dinding ini hanya terdiri dari protoplasma yang dibungkus oleh membran sel yang disebut ‘protoplas’ atau ‘bentuk-L’ (L untuk Lister Institute di London).
Setelah lisozim dihilangkan, mereka dapat melanjutkan pertumbuhan dengan mensintesis dinding sel mereka. Jika protoplas mempertahankan potongan dinding sel, sehingga sel mengambil bentuk amoeboid, menonjol keluar dimana tidak ada dinding sel, itu disebut ‘spheroplast’.
Spheroplast juga terbentuk, ketika peptidoglikan dinding sel gram negatif dihilangkan dengan pengobatan dengan lisozim. Di sini, protoplasma diapit oleh dua lapisan, membran sel dan lapisan LPS.
Fungsi Dinding Sel:
(a) Bentuk proteksi Lisis:
Ini melindungi sel bakteri dari lisis osmotik, karena kebanyakan dari mereka tetap berada di lingkungan hipotonik. Hal ini karena; konsentrasi zat terlarut terlarut di dalam sel bakteri jauh lebih tinggi daripada di lingkungan. Ini mengembangkan tekanan turgor yang cukup besar di dalam sel, yang mungkin sebanding dengan tekanan di dalam ban mobil. Dinding sel membantu menahan tekanan ini.
(b) Bentuk dan Kekakuan:
Ini memberikan bentuk dan kekakuan pada sel bakteri.
9. Ruang Periplasma:
Ini adalah ruang tipis antara dinding sel dan membran sel (Gambar 2.10 dan 2.11). Itu diisi dengan plasma yang disebut periplasma. Periplasma mengandung beberapa protein, tiga di antaranya penting.
Mereka adalah, 1) enzim hidrolitik, yang membantu dalam degradasi awal molekul makanan bakteri, 2) protein pengikat, yang memulai proses pengangkutan substrat dan 3) kemoreseptor, yang terlibat dalam respons kemotaksis bakteri. Chemotaxis berarti gerakan menuju atau menjauh dari bahan kimia.
10. Membran Sel:
Ini adalah membran tipis di bawah dinding sel yang membungkus protoplasma sepenuhnya. Ini juga disebut ‘membran sitoplasma’ atau ‘membran plasma’. Ini adalah bilayer fosfolipid tertanam protein (Gambar 2.11).
Dua jenis protein tertanam dalam lapisan ganda. Salah satu jenis protein bertahan dengan kuat di lapisan ganda dan disebut ‘protein integral’. Jenis protein lainnya berlabuh secara longgar pada permukaan bilayer dan disebut ‘protein periferal’.
Komponen membran tidak statis; melainkan mereka dinamis dan selalu dalam keadaan bergerak seperti ‘cairan’. Protein memberikan penampilan ‘mosaik’ pada membran.
Itulah mengapa; membran unit biasanya dinyatakan melalui ‘model mosaik cair’. Bilayer fosfolipid terdiri dari dua lapisan yang saling berhadapan yang masing-masing terdiri dari satu lapisan molekul fosfolipid. Setiap molekul fosfolipid memiliki kepala dan sepasang ekor yang muncul darinya.
Kepala bermuatan dan hidrofilik (suka air), yang tetap berada di lingkungan berair. Ekornya tidak bermuatan dan hidrofobik (membenci air), sehingga mereka tetap tersembunyi di dalam membran jauh dari lingkungan berair. Membran semacam itu disebut ‘membran unit’.
Sterol, yang ada di membran unit semua eukariota sebagai zat penguat, tidak ada di membran unit hampir semua bakteri. Namun, molekul yang mirip dengan sterol, yang disebut ‘humanoid’ ada di beberapa bakteri, yang memainkan peran yang mirip dengan sterol pada eukariota. Salah satu humanoid penting adalah ‘diploptene’, yang memiliki 30 atom karbon.
Fungsi Membran Sel:
(a) Penghalang Permeabilitas:
Bagian dalam sel (sitoplasma) terdiri dari larutan garam, gula, asam amino, vitamin, koenzim dan berbagai macam bahan larut lainnya. Sifat hidrofobik membran sel membuatnya & penghalang ketat.
Meskipun beberapa molekul hidrofobik kecil dapat melewati membran melalui difusi, molekul hidrofilik dan bermuatan (sebagian besar molekul yang larut dalam air) tidak mudah melewatinya. Molekul-molekul ini diangkut melewatinya dengan sistem pengangkutan khusus.
Bahkan zat sekecil ion hidrogen (H + ) tidak berdifusi melintasi membran sel. Satu-satunya molekul yang menembus membran adalah air, yang cukup kecil dan tidak bermuatan untuk melewati molekul fosfolipid.
Dengan demikian, membran sel adalah penghalang kritis yang memisahkan bagian dalam sel dari lingkungannya dan mencegah kebocoran pasif konstituen sitoplasma ke dalam atau ke luar sel. Jika membran rusak, integritas sel hancur, isi internal bocor ke lingkungan dan sel mati.
(b) Penghalang Selektif:
Membran sel juga merupakan penghalang yang sangat selektif. Protein spesifik, yang disebut ‘sistem pengangkut-membran’ yang ada di dalam dan di dalam membran sel memungkinkan dan memfasilitasi pengangkutan zat-zat tertentu melewatinya dengan transpor aktif dan pasif.
Secara struktural, sistem pengangkutan membran adalah protein yang terbuat dari 12 heliks alfa yang berputar bolak-balik melalui membran untuk membentuk saluran, melalui mana zat yang akan diangkut dibawa masuk atau keluar sel.
Protein ini dapat mengikat zat tertentu untuk diangkut dan memindahkannya ke dalam atau keluar sel melintasi membran sesuai kebutuhan. Ini memungkinkan sel bakteri untuk memusatkan zat tertentu di dalam sel dan mengeluarkan zat tertentu dari sel ke luar.
Ada tiga jenis protein transpor membran, seperti 1) Uniporter, yang mengangkut satu jenis molekul hanya dalam satu arah melintasi membran, 2) Symporters, yang mengangkut satu jenis molekul dan selalu mengangkut jenis molekul lain bersamanya dalam arah yang sama dan 3) Antiporter, yang mengangkut satu jenis molekul dan selalu mengangkut jenis molekul lain dalam arah yang berlawanan.
Transportasi zat dapat terjadi dalam tiga cara, seperti 1) Transportasi sederhana, di mana transportasi dilakukan melintasi membran oleh energi dari gaya gerak proton, 2) Translokasi kelompok, di mana zat yang diangkut diubah secara kimiawi selama perjalanan melintasi membran dan 3) Transportasi ABC (kaset pengikat ATP), di mana protein pengikat periplasma dalam periplasma secara khusus berikatan dengan substansi, protein rentang membran, yang melintasi membran mengangkutnya melintasi membran dan sitoplasma Protein penghidrolisis ATP yang ada dalam sitoplasma memasok energi untuk peristiwa transportasi.
(c) Hidrolisis Ekstraseluler:
Membran sel mengeluarkan banyak enzim ke luar untuk memecah partikel makanan besar menjadi molekul ekstraseluler, yang dapat masuk ke dalam sel melalui membran.
(d) Respirasi:
Bakteri tidak memiliki mitokondria. Oleh karena itu, respirasi sel untuk mendapatkan energi dari bahan makanan dilakukan di membran sel yang mengandung enzim rantai pernapasan. Ini adalah perangkat untuk konservasi energi dalam sel.
Membran dapat berada dalam bentuk ‘bermuatan’ yang penuh energi, di mana pemisahan proton (H + ) dari ion hidroksil (OH-) terjadi di seluruh permukaannya. Pemisahan muatan ini adalah bentuk energi metabolisme, analog dengan energi potensial yang ada dalam baterai yang terisi daya.
Keadaan energi membran, disebut sebagai ‘kekuatan motif proton’ (PMF), bertanggung jawab untuk menggerakkan banyak fungsi yang membutuhkan energi dalam sel, termasuk beberapa bentuk transportasi, motilitas dan biosintesis mata uang energi sel, ATP. .
(e) Fotosintesis:
Bakteri fototrofik melakukan fotosintesis oleh organel khusus yang disebut ‘kromatofor’ yang dibentuk oleh imajinasi membran sel. Organel ini mengandung enzim dan pigmen untuk fotosintesis.
(f) Penyerapan Nutrisi:
Nutrisi di sekitarnya langsung diserap ke dalam sel bakteri melalui membran sel. ‘Mesosom’ adalah invaginasi membran dari membran sel ke dalam sitoplasma, yang meningkatkan luas permukaan untuk penyerapan yang efisien.
(g) Reproduksi:
Bakteri berkembang biak dengan pembelahan biner. Setiap sel membelah menjadi dua sel anak. Dalam prosesnya, membran sel membentuk dinding sel baru di antara sel-sel anak yang membelah.
11. Sitoplasma:
Substansi seperti cairan kental, yang mengisi kompartemen di dalam membran sel, disebut sitoplasma (Gambar 2.10 dan 2.11). Ini adalah cairan yang terbuat dari air, enzim dan molekul kecil. Tersuspensi dalam cairan adalah daerah yang relatif transparan dari bahan kromosom yang disebut ‘nukleoid’, butiran ribosom dan inklusi sitoplasma, yang menyimpan energi.
12. Nukleoid:
Kumpulan massa DNA inilah yang membentuk kromosom bakteri (Gambar 2.12). Karena bakteri adalah prokariota, DNA mereka tidak terkurung dalam nukleus yang terikat membran, seperti pada eukariota.
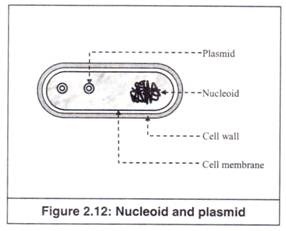
Namun, tidak tersebar di seluruh sel, melainkan cenderung berkumpul sebagai struktur yang berbeda dalam sel bakteri dan disebut ‘nukleoid’ (Gambar 2.12). Ribosom tidak ada di wilayah nukleoid, karena DNA nukleoid ada dalam bentuk seperti gel yang tidak termasuk materi partikulat.
Sebagian besar bakteri hanya mengandung satu kromosom, yang terdiri dari DNA untai ganda sirkular tunggal, meskipun beberapa bakteri memiliki DNA kromosom linier. Biasanya DNA kromosom dipelintir dan dilipat secara ekstensif menjadi ‘DNA super-gulungan’, untuk menampungnya di ruang kecil nukleoid.
DNA super-coiled lebih kompak daripada DNA sirkularisasi bebas. Bakteri hanya mengandung satu salinan dari setiap gen pada kromosomnya dan karenanya secara genetik ‘haploid’. Sebaliknya, eukariota mengandung dua salinan dari setiap gen dalam kromosomnya dan karenanya secara genetik diploid.
Gen dapat didefinisikan sebagai setiap segmen DNA, yang bertanggung jawab atas karakter tertentu dari organisme inang. Setiap karakter organisme ditentukan oleh gen tertentu, yang terdiri dari segmen DNA tertentu.
DNA kromosom bakteri mengandung ‘gen pemelihara rumah’ esensial, yang diperlukan untuk kelangsungan hidup dasar bakteri. DNA ini didistribusikan antara dua sel anak selama pembelahan sel.
Selain gen kromosom, ada juga gen non-kromosom yang ada di luar kromosom. Misalnya, plasmid mengandung gen spesifik, yang memberikan sifat khusus pada bakteri. Jumlah total dari semua gen yang ada dalam sel disebut ‘genom’.
13. Plasmid:
Ini adalah beberapa DNA non-kromosom yang ditemukan dalam sitoplasma beberapa bakteri (Gambar 2.12). Setiap plasmid terdiri dari sepotong kecil DNA beruntai ganda, yang dapat bereplikasi secara independen dari kromosom. Banyak plasmid agak kecil, tetapi ada juga yang cukup besar.
Namun, tidak ada plasmid yang sebesar kromosom. Plasmid tidak memiliki gen yang penting untuk pertumbuhan atau kelangsungan hidup bakteri. Mereka mungkin memiliki gen yang bertanggung jawab atas resistensi terhadap antibiotik, produksi toksin, atau produksi pelengkap permukaan yang penting untuk perlekatan dan pembentukan infeksi.
Jadi, plasmid mengandung gen yang memberikan sifat khusus pada bakteri, tidak seperti DNA kromosom, yang mengandung gen pemelihara rumah yang diperlukan untuk kelangsungan hidup dasar bakteri.
Beberapa plasmid, yang dapat berintegrasi ke dalam DNA kromosom inang disebut ‘episom’. Plasmid juga dapat dihilangkan dari sel bakteri inang dengan proses yang disebut ‘curing’. Beberapa plasmid banyak digunakan dalam manipulasi gen dan rekayasa genetika.
14. Ribosom:
Ini adalah badan granular yang ada di sitoplasma, di mana sintesis protein terjadi (Gambar 2.3). Jumlahnya sangat banyak sehingga keberadaannya membuat sitoplasma tampak gelap. Ukuran setiap ribosom bakteri adalah 70 S (70 Svedberg), yang terdiri dari dua subunit berukuran 50S dan 30S. Sebaliknya, ukuran setiap ribosom eukariotik adalah 80 S yang terdiri dari dua subunit berukuran 60S dan 40S.
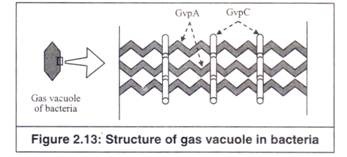
15. Vakuola Gas:
Vakuola gas adalah struktur berisi gas berbentuk spindel yang ditemukan di sitoplasma (Gambar 2.3 dan 2.13). Mereka adalah tabung silinder berongga, ditutup oleh tutup berbentuk kerucut berongga di setiap ujungnya. Jumlah mereka dalam sel dapat bervariasi dari beberapa hingga ratusan.
Panjang dan lebarnya bervariasi pada bakteri yang berbeda, tetapi vakuola dari setiap bakteri tertentu berukuran kurang lebih konstan. Mereka berongga, tapi kaku. Mereka tidak dapat ditembus air dan zat terlarut, tetapi dapat ditembus oleh sebagian besar gas. Oleh karena itu, komposisi gas di dalamnya sama dengan komposisi gas tempat organisme tersuspensi.
Gas hadir dalam vakuola sekitar 1 atm. tekanan. Oleh karena itu, kekakuan membran vakuola gas sangat penting bagi vakuola untuk menahan tekanan yang diberikan dari luar. Karena itu, ia terdiri dari protein yang mampu membentuk membran kaku daripada lipid, yang akan membentuk membran cair yang sangat mobile.
Ada dua jenis protein, yang membuat membran. Mereka adalah 1) GvpA dan 2) GvpC (protein vakuola gas A dan C). GvpA adalah protein vakuola gas utama, yang kecil dan sangat hidrofobik (Gambar 2.13). Ini adalah protein cangkang dan membentuk 97% dari total protein vakuola gas.
Vakuola gas terbuat dari beberapa molekul protein GvpA yang disejajarkan sebagai ‘tulang rusuk’ paralel yang membentuk permukaan kedap air. Protein GvpA terlipat sebagai (3 lembar dan dengan demikian, memberikan kekakuan yang cukup besar pada keseluruhan struktur vakuola. GvpC adalah protein yang lebih besar, tetapi hadir dalam jumlah yang jauh lebih kecil.
Ini memperkuat cangkang vakuola gas dengan bertindak sebagai penghubung silang, mengikat beberapa tulang rusuk GvpA menjadi satu seperti penjepit. Bentuk akhir dari vakuola gas, yang dapat bervariasi pada organisme yang berbeda dari panjang dan tipis hingga pendek dan gemuk adalah fungsi dari bagaimana protein GvpA dan GvpC disusun untuk membentuk vakuola utuh.
Karena vakuola gas diisi dengan gas, densitasnya sekitar 5-20% dari densitas sel sebenarnya. Dengan demikian, vakuola gas yang utuh menurunkan kepadatan sel bakteri, sehingga memberikan daya apung pada mereka.
Sebagian besar bakteri air terapung memiliki vakuola gas, yang memberikan daya apung dan membantu mereka mengapung di permukaan air atau di mana saja di kolom air. Vakuola gas juga bertindak sebagai sarana motilitas, memungkinkan sel mengapung naik turun di kolom air sebagai respons terhadap faktor lingkungan.
Misalnya, organisme fototropik akuatik mendapat manfaat dari vakuola gas, yang membantu mereka menyesuaikan posisinya dengan cepat di kolom air ke wilayah di mana intensitas cahaya untuk fotosintesis optimal. Membran vakuola gas tidak dapat menahan tekanan hidrostatik yang tinggi dan dapat runtuh, menyebabkan hilangnya daya apung. Setelah runtuh, vakuola gas tidak dapat dipompa kembali.
16. Inklusi:
Bahan cadangan tak hidup yang tersuspensi dalam sitoplasma disebut ‘inklusi’ (Gambar 2.3). Sebagian besar inklusi dibatasi oleh membran non-unit tipis yang terdiri dari lipid yang memisahkannya dari sitoplasma di sekitarnya.
Fungsi mereka adalah penyimpanan energi atau blok bangunan struktural. Bakteri yang berbeda mengandung berbagai jenis inklusi.
Beberapa inklusi umum yang ditemukan dalam sel bakteri dijelaskan di bawah ini:
(a) Butiran Poli-β-hidroksibutirat (PHB):
Ini adalah butiran yang terdiri dari poli- Asam β-hidroksibutirat (PHB), yang merupakan zat mirip lipid. Monomer asam β-hidroksibutirat terikat satu sama lain dalam rantai dengan ikatan ester untuk membentuk polimer poli- Asam β-hidroksibutirat. Polimer agregat menjadi butiran.
(b) Butiran Poli-β-hidroksialkanoat (PHA):
Ini adalah istilah kolektif yang digunakan untuk menggambarkan semua kelas polimer penyimpan karbon/energi yang ditemukan dalam sitoplasma bakteri. Butiran PHA yang paling umum adalah butiran PHB. Panjang monomer dalam polimer dapat bervariasi, dari sesingkat C 4 (dalam butiran PHB) hingga sepanjang C 18 (dalam butiran lain) di banyak bakteri.
(c) Butiran Volutin:
Ini adalah butiran yang terdiri dari polifosfat. Pewarna dasar, biru toluidin menjadi ungu kemerahan, bila bergabung dengan butiran polifosfat. Fenomena perubahan warna ini disebut ‘metachromasi’. Oleh karena itu, butiran volutin juga disebut ‘butiran metachromatic’.
(d) Butiran Glikogen:
Ini adalah polimer mirip pati yang terbuat dari subunit glukosa. Butiran glikogen biasanya lebih kecil dari butiran PHB. Ini adalah depot penyimpanan karbon dan energi dalam sel bakteri.
(e) Butiran Belerang:
Ini adalah butiran belerang unsur yang dihasilkan oleh akumulasi dalam sitoplasma. Unsur belerang berasal dari oksidasi senyawa belerang tereduksi seperti hidrogen sulfida dan tiosulfat. Butiran tetap ada selama sumber belerang tereduksi ada. Saat sumbernya menghilang, dimanfaatkan oleh sel bakteri, butiran juga menghilang perlahan.
(f) Magnetosom:
Magnetosom adalah butiran magnetit mineral besi, Fe 3 O 4 . Mereka menanamkan dipol magnet permanen ke sel bakteri, memungkinkannya merespons medan magnet. Sebuah membran yang mengandung fosfolipid, protein dan glikoprotein mengelilingi setiap magnetosom.
Protein membran berperan dalam presipitasi Fe 3+ (dibawa ke dalam sel dalam bentuk terlarut oleh agen pengkhelat) sebagai Fe 3 0 4 dalam magnetosom yang sedang berkembang. Morfologi magnetosom tampaknya spesifik spesies, bervariasi dalam bentuk dari persegi ke persegi panjang hingga berbentuk paku pada bakteri tertentu.
Magnetosom terutama ditemukan pada bakteri air. Bakteri yang menghasilkan magnetosom berorientasi dan bermigrasi di sepanjang medan geomagnetik. Fenomena ini disebut ‘magnetotaxis’. Bakteri, yang menunjukkan magnetotaxis, disebut ‘bakteri magnetotactic’.
Tidak ada bukti bahwa bakteri magnetotaktik menggunakan sistem sensorik bakteri kemotaktik atau fototaktik. Alih-alih, penyelarasan magnetosom dalam sel bakteri hanya memberikan sifat magnetik padanya, yang kemudian mengarahkan sel ke arah tertentu di lingkungannya. Jadi, istilah yang lebih baik untuk menggambarkan organisme ini adalah ‘bakteri magnetik’.
17. Endospora:
Ini adalah sel terdiferensiasi yang terbentuk di dalam sel bakteri berbentuk batang gram positif tertentu, yang dapat menahan kondisi buruk yang parah seperti panas, dingin, radiasi, penuaan, bahan kimia beracun, dan kekurangan nutrisi.
Sebagian besar, tiga genera bakteri berbentuk batang berikut dapat menghasilkan endospora dalam kondisi buruk:
- Bakteri berbentuk batang aerobik
- Bacillus spp.
- Bakteri berbentuk batang anaerobik
- Clostridium spp.
3.Desulfotomaculum spp.
Struktur Endospora:
Struktur endospora jauh lebih kompleks daripada struktur sel vegetatif karena memiliki banyak lapisan. Lapisan dari luar ke dalam diberikan di bawah ini (Gambar 2.14).
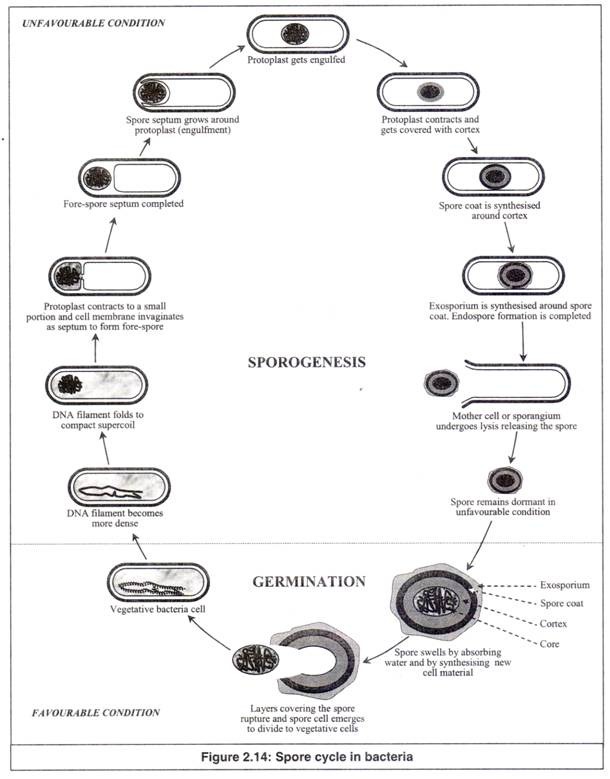
(sebuah) Eksosporium:
Ini adalah lapisan terluar, yang merupakan penutup tipis dan halus, terbuat dari protein.
(b) Lapisan spora:
Itu terbuat dari lapisan protein khusus spora. Ini bertindak seperti saringan yang mengeluarkan molekul beracun besar seperti lisozim, tahan terhadap banyak molekul beracun dan mungkin juga mengandung enzim yang terlibat dalam perkecambahan spora.
(c) Korteks:
Itu terbuat dari peptidoglikan yang terhubung secara longgar.
(d) Inti:
Ini berisi struktur sel bakteri yang biasa, yang meliputi dinding sel (dinding inti), yang menutupi protoplas spora. Protoplas spora terdiri dari membran sel, sitoplasma, nukleoid, ribosom, inklusi dan sebagainya.
Dengan demikian, spora berbeda secara struktural dari sel vegetatif terutama dalam jenis struktur yang ditemukan di luar dinding inti.
Properti Inti Endospora:
Inti dari endospora dewasa sangat berbeda dari sel vegetatif, dari mana ia terbentuk.
Sifat-sifat inti endospora adalah sebagai berikut:
(a) Salah satu zat kimia yang merupakan ciri khas endospora, tetapi tidak terdapat dalam sel vegetatif adalah asam dipikolinat (DPA). Itu terletak di inti.
(b) Spora juga mengandung ion kalsium yang tinggi, sebagian besar bergabung dengan asam dipikolinat. Kompleks asam kalsium-dipicolinic inti mewakili sekitar 10% dari berat kering endospora.
(c) Selain memiliki kandungan asam kalsium-dipikolinat (Ca-DPA) yang melimpah, inti berada dalam keadaan dehidrasi sebagian. Inti dari endospora dewasa hanya mengandung 10-30% kandungan air sel vegetatif dan dengan demikian, konsistensi sitoplasma inti adalah gel.
Dehidrasi inti sangat meningkatkan ketahanan panas endospora serta memberikan ketahanan terhadap bahan kimia, seperti hidrogen peroksida (H 2 O 2 ) dan menyebabkan enzim yang ada di inti tetap tidak aktif.
(d) Selain kandungan air spora yang rendah, pH sitoplasma inti sekitar satu unit lebih rendah daripada pH sel vegetatif.
(e) Inti mengandung protein spesifik inti tingkat tinggi yang disebut ‘protein spora larut asam kecil’ (SASPs). Ini disintesis selama proses spekulasi dan memiliki dua fungsi. Pertama, mereka mengikat erat DNA di inti dan melindunginya dari potensi kerusakan akibat radiasi UV, pengeringan dan panas kering. Kedua, mereka berfungsi sebagai sumber karbon dan energi untuk pertumbuhan sel vegetatif baru melalui proses perkecambahan.
Siklus Spora:
Dalam kondisi buruk, spora diproduksi di dalam sel vegetatif melalui dehidrasi dan kontraksi isi sel (Gambar 2.14). Spora ini, terbentuk di dalam sel bakteri, disebut ‘endospora’. Jika kondisi buruk memburuk, sel pecah melepaskan endospora, yang sekarang menjadi sel dorman independen yang disebut ‘spora’.
Proses dimana sel vegetatif menghasilkan spora, disebut ‘sporogenesis’ atau ‘sporulasi’. Spora tetap dalam bentuk dorman yang tidak aktif sampai kondisi buruk tetap ada. Setelah kembali ke kondisi yang menguntungkan, spora kembali ke bentuk vegetatif dan menjadi aktif kembali.
Lapisan yang menutupi spora pecah dan spora memunculkan sel vegetatif yang aktif secara metabolik. Proses, dimana spora berubah menjadi sel vegetatif, disebut ‘perkecambahan’.
Proses siklik perubahan dari bentuk vegetatif menjadi spora dalam kondisi yang merugikan dan kembali ke bentuk vegetatif dalam kondisi yang menguntungkan disebut ‘siklus spora’.
Dua bagian dari siklus spora diberikan di bawah ini:
- Sporulasi:
Sporulasi melibatkan serangkaian peristiwa yang sangat kompleks dalam diferensiasi seluler (Gambar 2.14). Banyak perubahan yang diarahkan secara genetis dalam sel mendasari konversi dari sel yang lembab dan aktif secara metabolik menjadi spora yang kering, tidak aktif secara metabolik tetapi sangat resisten.
Perubahan struktural yang terjadi pada sel bersporulasi dapat dibagi menjadi beberapa tahap. Seluruh proses sporulasi memakan waktu sekitar beberapa jam. Studi genetik telah menunjukkan bahwa sebanyak 200 gen terlibat dalam proses sporulasi yang khas.
Selama sporulasi, sintesis beberapa protein yang terlibat dalam fungsi sel vegetatif berhenti dan sintesis protein spora spesifik dimulai. Protein ini disebut ‘protein spora larut asam kecil’ (SASPs).
Ini dicapai dengan aktivasi berbagai gen spesifik spora termasuk spo, ssp (yang mengkodekan SASP) dan banyak gen lain sebagai respons terhadap kondisi lingkungan yang merugikan. Protein yang dikodekan oleh gen-gen ini mengkatalisasi rangkaian peristiwa yang mengubah sel vegetatif menjadi spora.
Spora mendapatkan ketahanan panasnya melalui beberapa mekanisme, yang belum dijelaskan dengan jelas. Namun, jelas bahwa lapisan kedap air, seperti korteks spora dan mantel spora melindungi spora dari kerusakan fisik.
Faktor utama yang mempengaruhi ketahanan panas suatu spora adalah jumlah dan keadaan air di dalamnya. Selama sporulasi, protoplasma berkurang ke volume minimum, sebagai akibat dari akumulasi Ca 2+ , protein spora kecil yang larut dalam asam (SASP) dan sintesis asam dipicolinic, yang mengarah pada pembentukan struktur seperti gel.
Pada tahap ini, korteks tebal terbentuk di sekitar inti protoplas. Kontraksi korteks menghasilkan protoplas yang menyusut dan dehidrasi dengan kandungan air hanya 10-30% dari sel vegetatif. Kandungan air protoplas dan konsentrasi SASP menentukan ketahanan panas spora.
Jika spora memiliki konsentrasi SASP yang rendah dan kandungan air yang tinggi, spora tersebut menunjukkan ketahanan panas yang rendah. Jika memiliki konsentrasi SASP yang tinggi dan kadar air yang rendah, ini menunjukkan ketahanan panas yang tinggi. Air bergerak bebas masuk dan keluar dari spora. Jadi, bukan impermeabilitas lapisan spora, melainkan bahan seperti gel dalam protoplas spora, yang tidak memasukkan air.
- Perkecambahan:
Endospora mungkin tetap tidak aktif selama bertahun-tahun, tetapi dapat berubah kembali menjadi sel vegetatif dengan relatif cepat. Proses ini melibatkan tiga langkah; aktivasi, perkecambahan dan pertumbuhan. ‘Aktivasi’ melibatkan pengaktifan spora dan membuatnya siap untuk perkecambahan.
‘Perkecambahan’ melibatkan hilangnya pembiasan mikroskopis spora dan hilangnya ketahanan terhadap panas dan bahan kimia. Spora kehilangan komponen asam kalsium-dipikolinat dan korteks. Selain itu, SASP terdegradasi.
‘Perkembangan’ melibatkan pembengkakan yang terlihat karena penyerapan air serta sintesis DNA, RNA, dan protein baru. Sel muncul dari selubung spora yang rusak dan akhirnya mulai membelah. Sel kemudian tetap dalam pertumbuhan vegetatif sampai kondisi lingkungan mendukung.
Saat mensterilkan bahan untuk membuatnya bebas bakteri, bakteri yang ada di dalamnya dibunuh dengan beberapa cara. Namun, jika bahan tersebut mengandung pembentuk spora, sulit untuk mensterilkannya. Semakin banyak jumlah spora dalam bahan, semakin tinggi tekanan yang diperlukan untuk mensterilkannya.
Untuk koloni bakteri pembentuk spora yang tumbuh pada pelat agar atau miring, seiring bertambahnya usia bakteri, mereka membentuk spora karena penuaan, defisiensi nutrisi, dan akumulasi metabolit.



