Baca artikel ini untuk mempelajari tentang struktur sel hewan!
Sel hewan dari unit struktural dasar semua jaringan dan organ tubuh. Tubuh kita memulai keberadaannya saat pembuahan dari satu sel, zigot diploid. Yang terakhir, melalui serangkaian proses dalam kehidupan sebelum dan sesudah kelahiran – pembelahan sel, pertumbuhan, diferensiasi, kematian sel terprogram (apoptosis) dan pematangan – pada akhirnya diubah menjadi manusia dewasa yang matang. Oleh karena itu, penting untuk mempertimbangkan anatomi seluler mikroskopis sejak awal.
Sumber Gambar : cdn.theatlantic.com/newsroom/img/posts/cells.jpg
Secara umum, sel adalah massa protoplasma yang mengandung nukleus. Sel darah merah mamalia, sebaliknya, tidak berinti dan memiliki masa hidup sekitar 120 hari. Beberapa sel dalam sumsum tulang merah berinti banyak. Protoplasma sel adalah emulsi heterogen dalam fase air yang mengandung zat kimia kompleks untuk proses metabolisme dan untuk penyimpanan bahan herediter.
Dalam bentuk kehidupan yang lebih rendah, seperti bakteri dan beberapa alga, sel tidak memiliki nukleus dan materi herediter dan metabolik tidak terpisah satu sama lain. Kelompok organisme yang memiliki sel tidak berinti ini dikenal sebagai prokariota.
Dalam bentuk kehidupan yang kompleks (membentang dari amuba ke manusia), sel-selnya mengandung nukleus yang terikat membran di mana informasi herediter disimpan dalam DNA kromosom, dan komponen sel lainnya di luar nukleus dikenal sebagai sitoplasma. Organisme seperti itu yang mengandung sel berinti disebut eukariota. Oleh karena itu, protoplasma digunakan dalam arti yang lebih luas, termasuk nukleus dan sitoplasma.
Pada hewan berukuran besar, ukuran selnya tidak bertambah tetapi jumlahnya bertambah. Karena ketika sitoplasma meningkat dalam jumlah yang cukup, DNA inti (gen) tidak dapat mengatur proses metabolisme sel, dan pada saat yang sama nutrisi sel dengan difusi menderita dari pinggiran ke pusat. Dengan demikian pembelahan sel adalah pilihan terbaik Alam untuk memulihkan hubungan yang optimal antara nukleus dan sitoplasma. Dalam sel normal, rasio inti-sitoplasma adalah sekitar 1:4 sampai 1:6. Tetapi pada sel ganas, nuklei berukuran besar secara tidak proporsional untuk sel, dan rasio inti-sitoplasma dapat mendekati 1:1
Struktur Sel Hewan:
Setiap sel terdiri dari membran sel, nukleus dan sitoplasma. Sel bervariasi dalam bentuk dan ukuran. Bentuknya mungkin pipih, kubik, kolumnar, fusiform, stellate, piramidal, berbentuk labu dan sebagainya. Ukuran sel bervariasi dari sekitar 5 µm sampai 50 µm. Sel telur manusia yang matang adalah salah satu sel terbesar, berukuran sekitar 130 malam.
Membran Sel:
Batas luar sel dikenal sebagai membran sel atau membran plasma. Ini semipermeabel dan mikroskop elektron mengungkapkan bahwa itu terdiri dari tiga lapisan yang ditumpangkan: lapisan luar terdiri dari protein, lapisan perantara fosfolipid bimolekuler, dan lapisan dalam protein. Ketebalan total membran sekitar 75A.
Lapisan protein memberikan elastisitas dan ketahanan mekanis relatif pada sel, dan lapisan fosfolipid memberikan permeabilitas pada bahan-bahan yang larut dalam lipid. Lapisan protein luar tebal sekitar 25A, dan ditutupi oleh lapisan sel yang dikenal sebagai glikokaliks yang terdiri dari tulang punggung glikoprotein yang mendukung asam sialat bermuatan negatif sebagai rantai samping terminal. Banyak antigen jaringan, termasuk antigen histokompatibilitas utama (MHC), terletak di lapisan sel.
Kompleks selubung sel membran plasma memberikan gaya elektrostatik untuk mengikat sel yang identik untuk membentuk jaringan spesifik dan membantu dalam transpor aktif ion Na + dan K + melintasi membran. Lapisan lipid menengah setebal 25A hingga 35A terdiri dari dua baris molekul fosfolipid, ujung kepala setiap molekul larut dalam air (hy drophilic) dan menghadap ke lapisan protein, dan ujung molekul lainnya tidak larut dalam air ( hidrofobik) dan bertemu satu sama lain di tengah membran.
bagian dalam tebalnya sekitar 25A dan agak berbeda dari lapisan protein bagian luar, karena tidak memiliki lapisan sel. Model membran sel trilaminar ini dikenal sebagai membran unit, karena ditemukan di sebagian besar organel yang terikat membran intraseluler. Bukti terbaru menunjukkan model mozaik cair dari membran plasma, di mana protein tertanam atau mengambang pada kedalaman yang bervariasi dalam lapisan ganda lipid (Gambar 2.1). Beberapa protein melintasi seluruh tugas dari dalam sel. Ini membantu menjelaskan bahwa berbagai protein permukaan reseptor secara aktif dipindahkan pada permukaan sel oleh elemen sitoskeletal internal.
Lapisan protein membran plasma relatif kaya akan asam amino, asam glutamat. Beberapa molekul protein dari lapisan luar terhubung dengan polisakarida bercabang , residu terminal yang bermuatan negatif asam sialat. Selaput plasma dari sel yang berdekatan dari jaringan padat (mis. Epitel) biasanya dipisahkan oleh celah sekitar 20 nm; jarak tersebut mungkin disebabkan oleh tolakan elektrostatik, tetapi gaya perekat untuk mengikat sel dibantu oleh selubung sel dan adanya divalen Ca ++ . Protein di zona hidrofobik dari lipid bimolekuler relatif kaya akan asam amino, leusin.
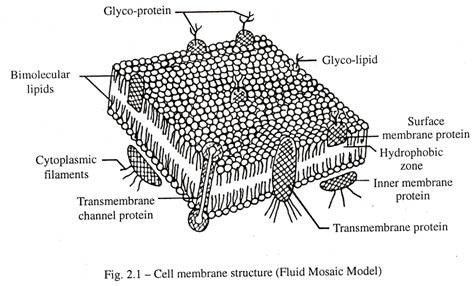
Beberapa molekul protein dari ketebalan bagian dalam membran sebagai protein membran trans yang mengandung saluran untuk difusi , sementara yang lain hanya menembus sebagian melalui itu. Bilayer fosfolipid dalam keadaan cair dan memungkinkan pergerakan protein di sepanjang bidang membran kecuali jika mereka adalah lapisan membran plasma memberikan jangkar ke mikrofilamen dan mikrotubulus, yang bertindak sebagai sitoskeleton untuk mengubah bentuk sel atau memberikan gerakan. dari sel.
Teknik freeze-fracture etching (FFE) mengungkapkan detail baru dari struktur membran. (Gbr. 2.2)
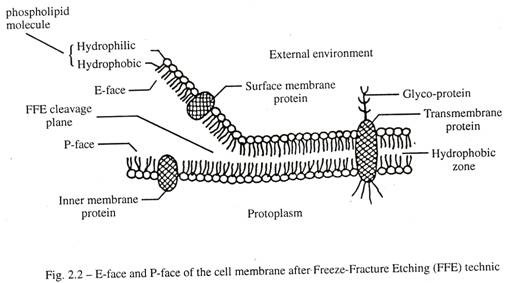
- Bidang fraktur melewati antara selebaran bagian dalam dan luar membran plasma melalui zona hidrofobik fosfolipid bimolekuler. Komponen lipid sebagian besar terdiri dari fosfatidil kolin dan kolesterol.
- Selebaran yang menghadap ke lingkungan luar disebut E-face.
- Selebaran yang menghadap protoplasma disebut P-face.
Kadang-kadang membran sel antara sel-sel yang berdekatan menghilang selama pembelahan sel , dan massa multinuklear yang disebut syncytium terbentuk. Lapisan protein dan lipid alternatif dari membran sel diwakili oleh selubung mielin saraf perifer karena spiral mesaxon (berasal dari membran sel) sel Schwann di sekitar serat saraf individu.
Fungsi membran plasma:
- Mempertahankan bentuk sel dan menyediakan lingkungan mikro untuk fungsi sel.
- Permeabilitas membran
saya. Hal ini memungkinkan lewatnya air dan gas seperti 0 2 dan C0 2 secara bebas, karena mereka sangat larut dalam lapisan ganda lipid.
- Zat yang larut dalam lemak seperti hormon steroid dapat masuk ke sitoplasma melalui lapisan lipid bimolekuler tanpa melewati saluran protein.
aku aku aku. Membran sel praktis tidak permeabel terhadap protein intraseluler dan anion organik lainnya.
- Banyak saluran protein membran trans memungkinkan permeabilitas selektif untuk ion seperti natrium, kalium, kalsium, klorida, dan bikarbonat. Bagian molekul yang lebih kecil seperti glukosa, asam amino dan prekursor asam nukleat juga terjadi melalui rute tersebut. Setiap saluran khusus untuk satu jenis ion atau molekul.
Beberapa saluran terus terbuka (saluran bocor) sedangkan yang lain mungkin membuka atau menutup (saluran berpagar) [Gbr. 2.3(a)]. Saluran yang terjaga keamanannya dapat terbuka akibat perubahan tegangan membran (voltage-gated), atau setelah berikatan dengan bahan kimia (ligÂand-gated). Beberapa saluran terbuka saat membran diregangkan. Saluran bocor sering dimanfaatkan oleh K + . Na + melewati saluran voltage-gated, ketika tegangan membran diturunkan. Sebuah saluran ligand-gated khas adalah reseptor nikotinat asetilkolin.
Beberapa protein membran bertindak sebagai pembawa, karena mereka mengangkut bahan melintasi membran plasma dengan mengikat ion dan molekul lain dan mengubah konfigurasinya untuk transportasi. Molekul berpindah dari area dengan konsentrasi tinggi ke area dengan konsentrasi rendah, menuruni gradien kimiawi, dan kation berpindah ke area bermuatan negatif sedangkan anion berpindah ke area bermuatan positif, menuruni gradien listriknya.
Ketika protein pembawa memindahkan zat ke bawah gradien kimia atau listriknya, itu disebut difusi terfasilitasi di mana tidak ada input energi yang diperlukan. Transpor glukosa menuruni gradien konsentrasinya dari cairan ekstraseluler (EcF) ke sitoplasma sel adalah contoh tipikal dari difusi terfasilitasi.
Pembawa lain mengangkut zat melawan gradien kimia dan listriknya. Bentuk transpor ini membutuhkan energi dan disebut transpor aktif, dan pengangkutnya disebut pompa. Dalam hal demikian, energi disediakan oleh hidrolisis ATP dan molekul pembawa membawa enzim, ATPase. Salah satu contoh klasik adalah Na + K + – ATPase (jadi adenosine triphosphatase yang diaktifkan dium-potassium), yang juga dikenal sebagai pompa Na + -K + – (lihat nanti).
- Protein pembawa bekerja dalam tiga cara berbeda: simport uniport dan antiport [Gbr. 2.3(b)].
Seragam mengangkut hanya satu zat ke bawah gradien konsentrasi; transporter glukosa bertindak sebagai uniport.
Simport membawa dua zat sekaligus dalam arah yang sama, menuruni gradien konsentrasi satu, misalnya Na + dan glukosa. Ini adalah contoh difusi terfasilitasi dimana Na + dan glukosa diangkut bersama-sama dari lumen usus ke sel mukosa.
Antiports memungkinkan pergerakan dua zat dalam arah yang berlawanan.
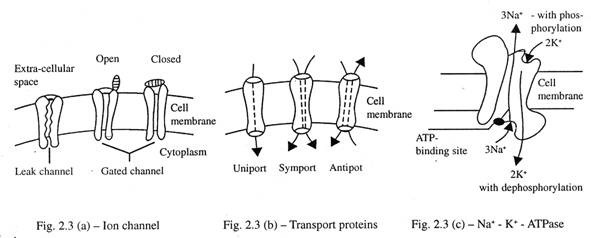
Aktivitas Na + -K + ATPase adalah contoh klasik dari antiport; itu bergerak 3Na + keluar dari sel sebagai ganti untuk setiap 2K + yang bergerak ke dalam sel ‘.
Na + -K + -ATPase-Ini adalah enzim dalam bentuk protein pembawa dan tertanam dalam membran plasma. Ini terdiri dari dua sub-unit, α dan β yang bervariasi dalam komposisi asam aminonya. Kedua sub-unit tersebut memiliki porsi intraseluler dan ekstraseluler.
Bagian intraseluler dari sub-unit berikatan dengan 3 Na + dan yang terakhir berikatan dengan ATP. Ini menghasilkan hidrolisis ATP menjadi ADP dan fosforilasi yang dihasilkan dari sub unit α menghasilkan perubahan konformasi yang terakhir; ini memungkinkan 3Na + untuk keluar dari sel. Sekarang, 2K + berikatan dengan bagian ekstraselular dari sub-unit yang kemudian mengalami defosforilasi dan kembali ke posisi semula, membawa 2K + pada saat yang sama ke dalam sel [Gambar 2.3©]
Potensi membran-Ada perbedaan potensial listrik antara sisi dalam dan luar membran sel, karena ion bermuatan listrik. Membran negatif di sisi dalam, dan positif di sisi luar. Sesuai kesepakatan, tanda minus ditulis untuk menunjukkan polaritas di sisi dalam membran. Di hampir semua sel hidup, potensial membran istirahat (RMP) berkisar antara -10mV hingga -90mV. Ini dapat diukur dengan meletakkan dua mikro-elektroda, satu di dalam dan satu lagi di luar membran sel, dan kemudian menghubungkannya ke osiloskop sinar katoda.
Ketika membran plasma sel saraf atau sel otot (dengan -70mV RMP) dirangsang dengan tepat, potensial istirahat turun menjadi sekitar -40mV hingga -50mV (depolarisasi) dengan muatan positif di dalamnya karena pembalikan kemampuan permeabilitas menjadi Na + dan K + .
Kejadian potensi membran- Dua protein transpor terutama bertanggung jawab atas potensi membran istirahat.
- a) Gradien konsentrasi untuk K + memfasilitasi difusinya keluar sel melalui saluran bocor untuk K + , tetapi gradien listriknya bekerja berlawanan arah. Akan tetapi, suatu kesetimbangan tercapai di mana kecenderungan K + untuk keluar dari sel diimbangi dengan kecenderungannya untuk berpindah ke dalam sel. Untuk mencapai kesetimbangan tersebut terdapat sedikit kelebihan kation di bagian luar dan anion di bagian dalam.
- b) Kondisi ini dipertahankan oleh Na + -K + – ATPase, yang memompa 3Na + keluar sel untuk setiap 2K + yang dipompanya di dalam sel. Influks Na + tidak mengkompensasi efluks K + , karena saluran kebocoran K + yang membuat membran lebih permeabel terhadap K + daripada Na + .
- Membran plasma bertindak sebagai permukaan sensorik dan membawa berbagai molekul reseptor, yang bergabung dengan molekul spesifik cairan jaringan dan mengubah aktivitas metabolisme sel melalui stimulasi atau penghambatan.
- Dari sekian banyak enzim yang ditanggung oleh membran sel, keberadaan adenilat siklase sangat mempengaruhi metabolisme sel. Stimulasi reseptor permukaan mengaktifkan adenil siklase yang bertindak sebagai pembawa pesan kedua dan menghasilkan peningkatan konsentrasi AMP siklik (adenosine monophosphate ) di dalam sel; yang terakhir menyebabkan perubahan sintesis DNA, ekspresi gen, sintesis protein dan peristiwa intraseluler lainnya. Sistem enzim serupa
mengontrol GMP siklik (guanidin monofosfat) yang memberikan efek antagonis dari AMP siklik. Beberapa hormon dan neuro-transmitter bekerja melalui pembawa pesan kedua.
Beberapa komponen fosfolipid membran sel (fosfoinositol) membantu dalam proses pengaturan kalsium di dalam sel dengan mengaktifkan fosfokinase dan fosforilasi berbagai komponen seluler.
- Pengenalan sel-sel identik dan perakitannya untuk membentuk jaringan spesifik dilayani oleh kompleks selubung sel membran plasma yang spesifik sel dan mengikat sel dengan kekuatan perekat.
- Membran plasma memiliki dua proses penting – endositosis dan ekstositosis.
Endositosis berarti penyerapan zat dari luar ke dalam sel dengan invaginasi lokal membran sel dalam bentuk vesikel endositik. Asupan cairan dengan metode ini dikenal sebagai pinositosis dan padatan seperti mikroorganisme dikenal sebagai fagositosis. Pada vesikel endositik lapisan dalam membran sel menjadi lapisan luar vesikel.
Eksositosis adalah proses pelepasan isi melalui vesikel sekretorik yang terikat membran dari dalam sel ke luar melalui fusi dengan membran plasma.
Nukleus:
Ini kurang lebih merupakan massa bulat yang ditutupi oleh amplop dan terletak di dalam sitoplasma di dekat pusat sel. Pada beberapa sel, nuklei berwajah terbuka dan menampilkan tampilan semi-transparan di mana isi nukleus divisualisasikan, sedangkan pada sel lain nukleus berwajah tertutup karena kondensasi bahan kromatin.
Ketika sebuah sel mati, nukleus menjadi piknotik dengan penyusutan dan menunjukkan massa hiperkromatin yang homogen. Nukleus diwarnai dengan pewarna dasar karena mengandung banyak DNA dan sejumlah kecil RNA. Inti terdiri dari: (a) selubung inti; (b) benang kromatin dalam sel istirahat, atau kromosom dalam sel pembagi; (c) nukleolus; (d) getah nuklir; (e) kromatin seks atau badan Barr.
Amplop nuklir [Gambar 2.4 (a), (b)]:
Ini menutupi nukleus dan terdiri dari dua unit membran (membran ganda) yang dipisahkan oleh cisterna perinuklear yang sempit. Membran luar bertabur ribosom dan sebenarnya berasal dari retikulum endoplasma kasar sitoplasma.
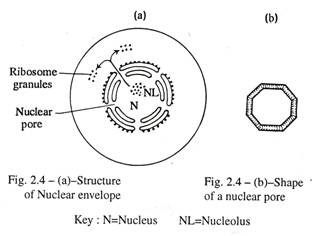
Membran bagian dalam adalah entitas yang terpisah dan bebas ribosom. Ini memberi keterikatan pada ujung kromosom dan lapisan padat kromatin selama antar fase. Banyak pori-pori inti berbentuk segi delapan hadir dalam selubung inti dan dibentuk oleh fusi membran inti luar dan dalam.
Setiap pori berdiameter sekitar 80 nm berbentuk corong, ujung luarnya lebih sempit daripada ujung dalam, dan bertindak sebagai diafragma untuk pertukaran nuklir-sitoplasma. Melalui pori-pori ini mRNA, rRNA, tRNA ditransmisikan dari nukleus ke sitoplasma, tetapi organel sitoplasma yang merusak seperti lisosom dicegah memasuki nukleus. Nukleus tipikal memiliki sekitar 3000-4000 pori.
Benang kromatin dan kromosom:
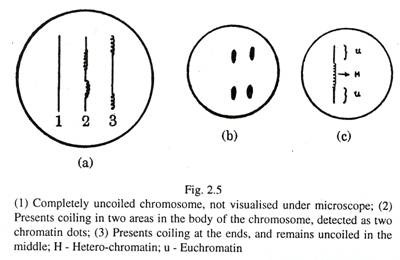
Pada fase istirahat atau interfase sel hewan [Gambar 2.5(a) dan (b)], nukleus mengandung jaringan benang atau butiran kromatin yang diwarnai dengan pewarna dasar. Kromosom individu tidak dapat diidentifikasi, karena selama interfase menjadi tidak berliku dan menipis. Di beberapa tempat kromosom masih tetap melingkar dan area ini divisualisasikan sebagai butiran atau titik kromatin. Oleh karena itu, butiran atau benang kromatin bukanlah segmen kromosom yang rusak. Segmen kromosom yang tidak digulung dikenal sebagai eukromatin, yang aktif secara genetik. Segmen melingkar kromosom disebut heterokromatin, yang secara genetik inert [Gambar. 2.5(c)].
Selama pembelahan sel, setiap kromosom menjadi lebih tebal, lebih pendek dan melingkar rapat sepanjang panjangnya. Karenanya kromosom individu divisualisasikan dan diidentifikasi. Kromosom adalah benang yang sangat ternoda dan jumlahnya konstan dalam suatu spesies. Pada manusia jumlahnya adalah 46 (diploid) pada semua sel somatik, tetapi 23 (haploid) pada sel benih dewasa.
46 kromosom disusun dalam 23 pasang; 22 pasang dikenal sebagai autosom yang mengatur karakter tubuh; pasangan yang tersisa dikenal sebagai kromosom seks atau gonosomes , yang terutama mengatur karakter seks. Satu anggota dari setiap pasangan berasal dari ayah, dan anggota lainnya berasal dari ibu. Pasangan terjadi antara kromosom yang identik, yang identik dalam panjang, posisi sentromer dan distribusi gen.
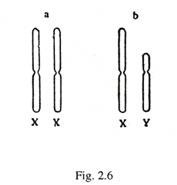
Kromosom berpasangan dikenal sebagai kromosom homolog. Pada wanita, kromosom seks memiliki panjang yang sama dan dilambangkan dengan XX [Gambar 2-6(a)]. Pada laki-laki, kromosom seks tidak sama panjangnya, dan dilambangkan dengan XY [Gambar. 2-6(b)]. Yang lebih panjang diwakili oleh X, dan yang lebih pendek oleh Y. Selama berpasangan keduanya memiliki bagian homolog dan non-homolog.
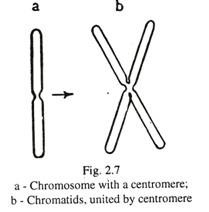
Setiap kromosom menghadirkan penyempitan yang dikenal sebagai sentromer atau kinetokor yang melekat pada gelendong akromatik selama pembelahan sel [Gambar. 2-7(a)]. Dalam profase pembelahan sel, setiap kromosom membelah secara longitudinal menjadi dua kromatid kecuali pada sentromer [Gambar. 2-7(b)].
Gen terletak di kromosom dalam seri linier. Gen adalah bagian dari molekul DNA spesifik dan mentransmisikan karakter yang diwariskan dari satu generasi ke generasi berikutnya. Gen juga bertanggung jawab untuk sintesis protein sel melalui messenger RNA, RNA ribosom, dan RNA transfer.
Nukleolus:
Ini adalah benda bulat yang sangat refraksi tanpa membran penutup, dan terletak dekat dengan membran nuklir [Lihat Gambar 2-4(a)]. Ini adalah massa terkompresi dari campuran butiran RNA (ribosom) dan protein. Sintesis RNA nukleolus diatur oleh gen-gen yang terletak pada penyempitan sekunder kromosom-kromosom yang memiliki badan satelit pada lengan pendeknya (anggota pasangan kromosom 13 sampai 15, 21 dan 22).
RNA dibebaskan dari nukleolus dan muncul di sitoplasma melalui pori-pori nuklir. Nukleolus menghilang selama profase dan muncul kembali selama telofase pembelahan sel.
Getah nuklir:
Ini adalah cairan yang mengandung protein yang mengisi celah antara benang kromatin dan membran inti. Ini berfungsi sebagai media untuk transportasi RNA ribosom dan RNA messenger ke pori-pori nuklir.
Kromatin seks atau badan Barr:
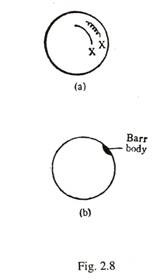
Selama interfase, badan cembung plano heterokromatin ditemukan di bawah membran nukleus pada wanita normal [ Gambar 2-8(b)], ini dikenal sebagai kromatin seks atau badan Barr. Selama pembelahan sel, badan Barr menghilang. Dari kromosom 2X pada wanita normal, salah satunya sangat melingkar dan anggota lainnya sangat tidak melingkar [Gambar. 2-8(a)].
Kromosom X yang tidak aktif secara genetik yang sangat melingkar membentuk tubuh Barr. Badan-badan ini membantu dalam sexing nuklir jaringan. Jumlah badan Barr dalam sel sama dengan jumlah total kromosom X dikurangi satu. Jadi pada wanita normal dengan kromosom 2X, jumlah tubuh Barr adalah satu; pada sindrom Triple X (XXX), jumlahnya meningkat menjadi dua.
Selama interfase, kromosom Y laki-laki menunjukkan di dalam nukleus massa yang sangat berfluoresensi yang dikenal sebagai F-body, ketika diwarnai dengan pewarna flurochrome dan dilihat di bawah mikroskop fluoresensi.
Sitoplasma:
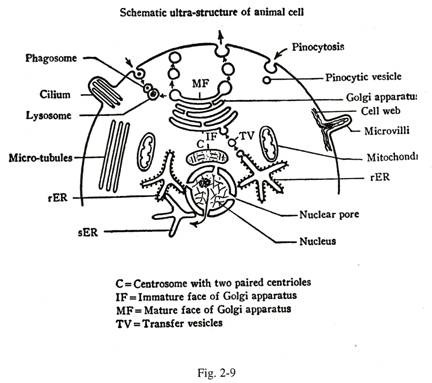
Ini adalah bagian dari protoplasma yang mengintervensi antara membran sel dan selubung inti. Sitoplasma atau sitosol terdiri dari dua bagian-organel atau elemen aktif, paraplasma atau inklusi seperti, glikogen, gumpalan lemak dan pigmen. Organel adalah sebagai berikut [Gambar. 2-9].
- Mitokondria;
- Butiran ribosom;
- Endoplasma
- Aparat Golgi; retikulum;
- Lisosom;
- Fagosom;
- Peroksisom;
- Sentriol dan mikrotubulus;
- Filamen dan serat;
Mitokondria:
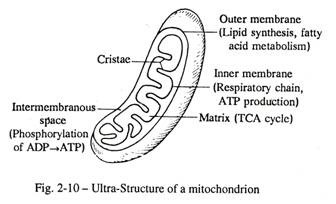
Setiap sel aktif menghadirkan banyak mitochondria yang berbentuk batang atau badan yang terikat membran vesikular. Badan-badan ini dapat dilihat di bawah mikroskop cahaya setelah pewarnaan dengan fuchsin asam atau pewarnaan supra-vital janus hijau. Mikroskop elektron mengungkapkan bahwa setiap mitokondria terdiri dari dua dinding membran, luar dan dalam, dipisahkan oleh ruang intermembran [Gambar. 2-9, 2-10]. Setiap dinding membran mewakili unit membran.
Membran bagian dalam dilipat membentuk partisi tidak lengkap yang dikenal sebagai krista mitokondria yang memberikan perlekatan pada enzim untuk fosforilasi ADP menjadi ATP oleh batang silinder. Bagian dalam setiap mitokondria diisi dengan cairan, matriks mitokondria, yang berisi DNA berbentuk lingkaran, RNA dan enzim pernapasan penting seperti bakteri. Oleh karena itu, dengan kemajuan evolusi, dinding bagian dalam mitokondria berasal dari bakteri yang dilemahkan, yang ditarik ke dalam sitoplasma sel hewan dan menjalani kehidupan simbiotik untuk menyelesaikan respirasi aerobik dari sel hewan yang diinvasi. Selain itu, mitokondria membelah dengan pembelahan yang mirip dengan bakteri.
Tiga enzim penting ditemukan di mitokondria: –
(a) Enzim siklus asam sitrat Kreb;
(b) Flavo-protein, dehydrogenase dan cytochrome yang merupakan enzim respirasi;
(c) Fosforilase oksidatif.
Fungsi:
- Mitokondria menyelesaikan respirasi sel dengan jalur aerobik dan menghasilkan energi tinggi melalui pembentukan ATP.
Gula dalam matriks sitoplasma mengalami degradasi tanpa bantuan oksigen (anaerobik) melalui proses glikolisis dan diubah menjadi asetil-koenzim A yang kemudian masuk ke dalam mitokondria, dimana asetil-koA bergabung dengan oksalo-asetat membentuk asam sitrat. Enzim siklus asam sitrat melalui beberapa reaksi dekarboksilasi menghasilkan Co 2 dan dengan bantuan dehidrogenase spesifik melepaskan empat pasang ion H + . Enzim pernapasan, flavo-protein dan cytochrome , kemudian mentransfer ion hidrogen keluar dari mitokondria hingga bergabung dengan oksigen dan membentuk air.
- Energi yang dibebaskan selama transpor ion hidrogen digunakan oleh fosforilase oksidatif untuk regenerasi ATP dari ADP dan fosfat anorganik. ATP kaya energi yang terbentuk di dalam mitokondria diambil oleh sitoplasma dalam kondisi aerob, sehingga 36 molekul ATP terbentuk per molekul glukosa. Ini adalah 18 kali energi yang diperoleh dalam keadaan anaerobik dari jalur glikolitik. Jadi mitokondria bertindak sebagai pembangkit tenaga sel.
- Bentuk melingkar DNA mungkin merupakan faktor pewarisan sitoplasma. Semua mitochondria berasal dari ibu. DNA mitokondria abnormal dapat menyebabkan kelemahan otot dan penyakit degeneratif SSP karena kegagalan metabolisme oksidatif. Ini dikenal sebagai sindrom sitopati mitokondria.
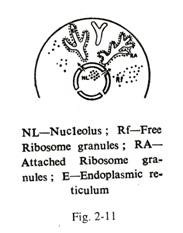
Butiran ribosom:
Granula ribosom terdiri dari RNA ribosom dan protein. Ribosom pada awalnya dikumpulkan dalam nukleolus dan disintesis oleh pengatur nukleolar dari satkromosom (kromosom 13, 14, 15, 21, 22 yang memiliki badan satelit). Dari nukleolus ribosom muncul di sitoplasma
melalui pori-pori inti (Gbr. 2-9, 2-11).
Di dalam sitoplasma beberapa ribosom tetap bebas, sementara yang lain melekat pada retikulum endoplasma yang membuat permukaannya kasar. Ribosom bebas membuat sitoplasma basofilik. Dalam sel embrionik dan sel ganas ribosom bebas berlimpah.
Pada eukariota setiap granula ribosom terdiri dari dua sub-unit, 40S dan 60S. S adalah singkatan dari satuan laju sedimentasi Svedberg. Rantai polinukleotida messenger RNA melewati sub-unit 40S dari ribosom yang memperlihatkan kodon triplet.
Sub unit 60S adalah tempat berlangsungnya sintesis protein melalui hubungan linier asam amino dengan bantuan antikodon RNA transfer (Gambar 2.12). Oleh karena itu, ribosom bebas mensintesis protein yang digunakan untuk metabolisme sel dan pertumbuhannya sendiri.
Kadang-kadang sejumlah ribosom dilampirkan ke rantai tunggal RNA kurir. Fenomena seperti itu dikenal sebagai poliribosom atau polisom.
Ini adalah sistem vesikel atau tubulus membran yang saling berkomunikasi, yang dapat meluas dari membran nuklir ke membran sel. Retikulum endoplasma atau ER memiliki dua jenis – kasar dan halus (Gbr. 2-9).
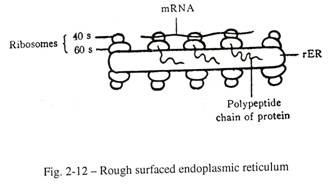
ER permukaan kasar, segera disebut rER memberikan perlekatan butiran ribosom ke permukaan luar vesikel membran dan menghasilkan kekasaran retikulum. Sub-unit yang lebih besar dari ribosom (60S) melekat pada permukaan RE dan sub-unit yang lebih kecil (40S) terletak pada matriks sitoplasma.
Rantai polipeptida dari protein yang disintesis dalam sub-unit yang lebih besar didorong ke dalam retikulum, tempat makromolekul protein disimpan dan selanjutnya dikirim ke luar sel sebagai produk sekretori (Gbr. 2-12). Oleh karena itu rER membantu dalam sintesis protein dan penyimpanannya. RER hadir di semua sel sekretori, seperti sel asinar pankreas.
ER atau sER permukaan halus diatur dalam jaringan tubulus plexiform, dan permukaan luarnya tidak memiliki butiran ribosom. Beberapa sel, seperti sel hati, memiliki rER dan sER. Ser membantu dalam sintesis lipid dan steroid. Protein yang disintesis dari rER ditransfer ke sER, di mana kompleks lipo-protein terbentuk.
Bahan protein lipo tersebut dari sel hati disalurkan melalui aparatus Golgi dan permukaan sel ke darah. Sel-sel hati membantu detoksifikasi obat larut lipid tertentu melalui enzim hidroksilasi dari sER. Dipercayai bahwa awalnya rER terbentuk, yang kemudian diubah menjadi sER dengan kehilangan butiran ribosom. Retikulum sarkoplasma sel otot lurik adalah contoh dari sER.
Aparat Golgi:
Aparatus Golgi (Gbr. 2.9, 2.13)—Ia terdiri dari cisternae membranosa berpermukaan halus dan padat yang disusun dalam tumpukan empat sampai enam, bersama dengan kelompok vesikel kecil di sekitar permukaannya. Aparatus hadir di sebagian besar sel, tetapi menonjol di sel sekretori di mana ia mengintervensi antara rER dan membran sel.
Dalam pewarnaan HE, aparatus menampilkan area yang jelas; karenanya disebut citra Golgi negatif. Di bawah mikroskop elektron, aparatus Golgi klasik memiliki dua wajah – wajah yang belum matang atau cis dengan permukaan cembung yang diarahkan ke rER, dan wajah dewasa atau trans dengan permukaan cekung yang diarahkan ke membran sel. Selain cisternae yang rata, vesikel kecil jaringan cis-Golgi dan trans-Golgi membentuk bagian integral dari kompleks Golgi.
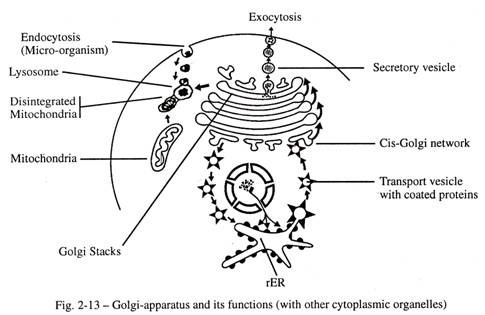
Wajah cis Golgi menerima vesikel trans port kecil dengan protein selubung khusus yang bertunas dari rER. Vesikel pengangkut membawa protein yang disintesis dari rER dan mengirimkan isinya ke cisterna pertama melalui fusi membran. Selama proses ini, vesikel pengangkut dicegat oleh vesikel penghubung jaringan cis-Golgi, yang memilih apakah protein sesuai untuk dikirim ke tumpukan Golgi; Namun, protein yang tidak tepat dikembalikan ke rER.
Di dalam aparatus Golgi bagian karbohidrat ditambahkan ke bahan protein dengan bantuan transferase dan glikoprotein terbentuk. Dari tepi cisterna pertama, protein yang dimodifikasi diangkut oleh tunas vesikuler dan kemudian fusi ke tepi cisterna berikutnya sampai cisterna terakhir tercapai di permukaan trans. Setelah serangkaian pemrosesan dan kondensasi, gelembung glikoprotein keluar dari permukaan trans aparatus Golgi sebagai vesikel yang menggelembung.
Penyortiran akhir protein yang dimodifikasi dan pengemasannya dalam bentuk vesikel dengan urutan asam amino terpilih terjadi di jaringan trans-Golgi. Yang terakhir menentukan tujuan paket vesikel; beberapa dipertahankan dalam sitoplasma sebagai lisosom, sementara yang lain keluar dari sel sebagai vesikel sekretorik dan mengirimkan isinya melalui membran sel melalui eksositosis. Selain sel sekretori, aparatus Golgi pada sel non sekretori membebaskan selubung sel (glikokaliks) di luar membran plasma. Kompleks membran sel-lapisan plasma mengerahkan gaya elektrostatik yang mengikat sel-sel identik untuk membentuk jaringan tertentu.
Lisosom:
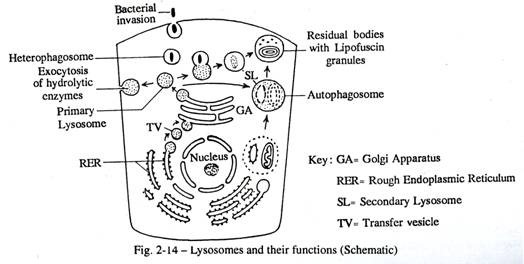
Lisosom merupakan vesikel membran berdinding tebal yang mengandung enzim hidrolitik yaitu protease, lipase dan asam fosfatase. Enzim ini, ketika dibebaskan dari lisosom, dapat mencerna zat tertentu yang berasal dari dalam sitoplasma atau dimasukkan ke dalam sel dari luar. Sebelum sel mati karena kekurangan oksigen atau karena alasan lain, lisosom bertindak sebagai vesikel autofagik dan menghancurkan semua organel sitoplasma. Oleh karena itu, mereka dikenal sebagai ‘kantong bunuh diri’ dari sel.
Setelah kematian, kecuali hewan difiksasi dengan fiksatif, lisosom pecah dan autolisis terjadi. Pada sel sehat, lisosom berfungsi melindungi dan menghancurkan penyerang bakteri tertentu (Gbr. 2-8, 2-3) bertindak sebagai vesikel heterofagik. Enzim lisosom mendegradasi berbagai zat yang merugikan di dalam sel. Ketiadaan kongenital dari enzim lisosomal tertentu menyebabkan akumulasi substratnya di dalam sel, menghasilkan penyakit penyimpanan, misalnya penyakit Tay-sachs, penyakit Gaucher. Lisosom seperti itu berlimpah dalam sel makrofag dan leukosit granular.
Lisosom kaya akan glikoprotein dan berasal dari permukaan aparatus Golgi yang matang sebagai lisosom primer. Dalam proses kerusakan sel, potongan mitokondria dan retikulum endoplasma yang tidak berfungsi bergabung dengan lisosom dan mengalami pencernaan. Badan yang menyatu seperti itu membentuk lisosom sekunder dan dikenal sebagai sitolisosom. Jadi lisosom diberkahi dengan fungsi menghilangkan sisa-sisa degradasi organel sitoplasma.
Beberapa sisa lisosom yang tidak larut setelah autophagy dan heterophagy membentuk badan residu yang dipertahankan secara permanen sebagai pigmen kepikunan yang terbuat dari lipofuscin kaya lipid (Gbr. 2-14). Pigmen ini ditemukan pada usia tua di dalam sistem saraf.
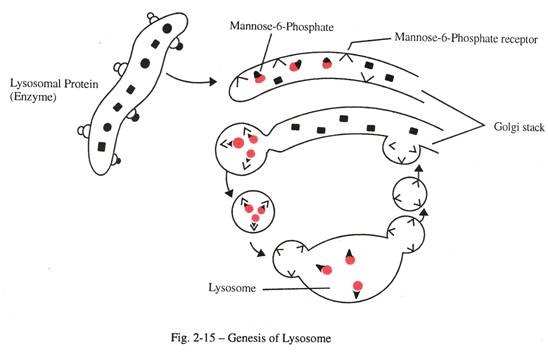
Genesis lisosom memerlukan modifikasi khusus protein di dalam aparatus Golgi (Gbr. 2-15). Ini melibatkan perlekatan manosa-6-fosfat ke beberapa rantai samping oligosakarida. Enzim lisosom mengikat residu manosa terfosforilasi ke reseptor spesifiknya yang ditemukan di permukaan dalam membran Golgi.
Setelah itu enzim yang terikat reseptor dicubit dari Golgi dan dilanjutkan untuk berfusi dengan lisosom. Setelah melepaskan protein sekretori dengan mannose-6-fosfat yang melekat pada lisosom, vesikel yang mengandung reseptor spesifik yang dimediasi enzim kembali ke Golgi untuk digunakan kembali. Keasaman isi lisosom (sekitar pH5) dipertahankan dengan memompa proton dari sitoplasma melalui transpor aktif.
fagosom:
Kadang partikel atau mikroorganisme hidup masuk ke dalam sitoplasma sel dari luar, ditutupi oleh lipatan membran sel. Vesikel membran seperti itu dikenal sebagai fagosom. Saat fagosom bersentuhan dengan lisosom, dinding umum di antara mereka menghilang dan enzim hidrolitik lisosom menghasilkan lisis bahan yang terkandung. Proses ini dikenal sebagai fagositosis, yang agak mirip dengan pinositosis. Dalam pinositosis, cairan masuk ke dalam sitoplasma, sedangkan dalam fagositosis, partikel padat berpartisipasi dalam proses tersebut.
Peroksisom:
Ini adalah vesikel kecil yang terikat membran dengan diameter sekitar 0,5-1,5 µm; karenanya disebut mikrobodi. Peroksisom hadir di sebagian besar sel berinti, dan lebih banyak di hepatosit dan sel tubulus ginjal. Mereka mengandung enzim oksidatif yang membantu detoksifikasi berbagai zat dan menghasilkan hidrogen peroksida; mereka juga berpartisipasi dalam (3-oksidasi rantai asam lemak. Kelebihan jumlah hidrogen peroksida dipecah oleh enzim, katalase.
Asal usul peroksisom itu aneh. Membran sel berasal dari multiplikasi peroksisom yang sudah ada sebelumnya, dan protein internalnya datang langsung dari sitosol melalui saluran pori membran plasmanya, melewati vesikel paket biasa dari rER dan aparatus Golgi.
Sentriol dan mikrotubulus:
Sentriol:
Setiap sel hewan, yang mampu membelah, memiliki dua sentriol di dalam sitoplasma dan dekat dengan membran inti. Daerah padat sitoplasma yang mengandung sentriol dikenal sebagai sentrosom. Setiap sentriol menghadirkan dua benda silinder yang ditempatkan pada sudut siku-siku satu sama lain. Dinding silinder menghadirkan sembilan bundel longitudinal dan setiap bundel terdiri dari tiga mikrotubulus yang tertanam dalam bahan fibrilar (Gbr. 2-16).
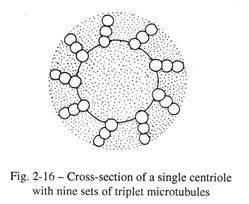
Sentriol membantu sintesis mikrotubulus gelendong achromatic selama pembelahan sel, dengan keterkaitan protein sitoplasma larut yang dikenal sebagai tubulin. Selama pembelahan sel, dua sentiol (masing-masing dengan dua badan silinder) dipisahkan satu sama lain oleh mikrotubulus yang tumbuh dari gelendong akromatik dan menempati kutub nukleus yang berlawanan (lihat Gambar 3-2). Mikrotubulus ini memanjang di antara sentriol yang berlawanan membentuk mikrotubulus kontinu dari gelendong. Pada metafase, membran nukleus menghilang dan mikrotubulus kromosom diatur dari protein tubulin oleh kinetokor, dua di antaranya berada di sisi sentromer setiap kromosom.
Mikrotubulus kromosom mendorong sentriol yang berlawanan lebih jauh sampai kromosom dengan kromatid berpasangan menempati ekuator gelendong. Dengan demikian gelendong akromatik sel mitosis terdiri dari mikrotubulus kontinu yang diatur oleh sentriol dan mikrotubul kromosom yang diatur oleh kinetokor. Masing-masing dari dua sel baru yang berasal dari pembelahan sel mengandung satu sentriol dengan dua badan silinder yang terletak tegak lurus satu sama lain.
Setelah itu sebuah sentriol terbentuk di dekat masing-masing sentriol lama, sehingga memulihkan komplemen normal sentriol.
Selain pembentukan gelendong, sentriol membantu tumbuhnya silia dan tubulus mikro bersama dengan proses perkembangan neuron.
Mikrotubulus:
Silia, flagela, dan sentriol terdiri dari mikrotubulus. Faktanya, semua sel hewan memiliki mikrotubulus yang dapat diatur atau tersebar. Mereka adalah struktur berfilamen yang terbuat dari protein tubulin terlarut. Mikrotubulus yang tersebar bertindak sebagai kerangka sel dan membantu pengangkutan berbagai zat termasuk makromolekul ke seluruh sitoplasma. Karena mikrotubulus terdiri dari protein kontraktil, mereka berkaitan dengan gerakan melalui silia, flagela dan oleh gelendong akromatik yang mendorong sentriol terpisah selama pembelahan sel.
Setidaknya tiga situs tersedia di sitoplasma yang bertindak sebagai pusat pengorganisasian mikrotubulus (MTOC) –
(a) Sentriol untuk mikrotubul kontinyu dari gelendong;
(b) Kinetokor kromosom untuk mikrotubulus kromosom;
(c) Badan basal silia untuk pertumbuhan mikrotubulus siliaris. Colchicine, suatu zat kimia, menghentikan penglihatan sel pada metafase dengan bergabung dengan protein tubulin dan mencegah pembentukan achromatic spindle.
Filamen dan fibril:
Ini adalah jaringan ultra-mikroskopik dari struktur berserabut yang berbeda dari mikrotubulus. Beberapa filamen lebih padat di bawah membran sel membentuk jaring sel. Filamen dan komponennya yang lebih tebal, fibril, bertindak sebagai pendukung internal sel. Beberapa filamen masuk ke inti pusat mikrovili, sementara yang lain membentuk filamen aktin dan miosin dari otot kontraktil.



