
Banyak komponen komplemen yang beredar secara fungsional tidak aktif. Aktivasi satu komponen komplemen menyebabkan aktivasi komponen komplemen kedua .
Komponen kedua yang diaktifkan bekerja pada komponen pelengkap ketiga; aktivasi komponen pelengkap lainnya berlanjut secara berurutan. Dengan demikian aktivasi sistem komplemen terjadi melalui mode seperti kaskade berurutan (misalnya mengatur 9 batu berdampingan dan mendorong batu pertama. Batu pertama jatuh di atas batu kedua dan mendorong batu kedua jatuh. Batu kedua jatuh di atas batu ketiga batu, yang jatuh pada batu ke-4 secara bergiliran, dan seterusnya sampai batu ke-9 yang terakhir jatuh.) (Gbr. 10.1).
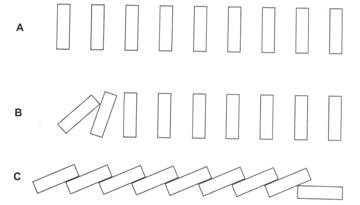
Gambar 10.1 A sampai C: Sebuah contoh untuk menggambarkan mode kaskade aktivasi komplemen.
(A) Sembilan batu disusun berdampingan. (B) Batu pertama didorong dan jatuh dan mendorong batu kedua. (C) Jatuhnya batu pertama pada akhirnya mengakibatkan jatuhnya batu kesembilan yang terakhir
Aktivasi komplemen terjadi melalui dua jalur yang berbeda:
saya. Jalur aktivasi komplemen yang ditemukan pertama kali disebut jalur klasik aktivasi komplemen.
- Jalur aktivasi komplemen yang ditemukan kemudian disebut jalur alternatif aktivasi komplemen. Mekanisme inisiasi aktivasi kedua jalur ini berbeda. Namun, kedua jalur tersebut mengarah pada pembelahan komponen komplemen 3 (C3). C3 adalah umum untuk kedua jalur dan peristiwa yang terjadi setelah pembelahan C3 serupa di kedua jalur.
1. Jalur Klasik Aktivasi Pelengkap:
Pengikatan antibodi terhadap antigen dalam sirkulasi atau antigen pada sel target (seperti mikroba) memulai aktivasi jalur komplemen klasik (Gambar 10.2). Pengikatan antibodi terhadap antigen mengekspos situs pengikatan C1q pada wilayah Fc molekul antibodi. Komponen pelengkap CI terbuat dari 3 protein yang ditunjuk C1q, C1r, dan Clqr2S2. Bagian C1q (dari CI) berikatan dengan situs pengikatan C1q pada antibodi yang terikat antigen.
sebuah†”
Pengikatan C1q ke antibodi menginduksi perubahan konformasi pada C1r. Perubahan konformasi membuat C1r menjadi enzim aktif, yang disebut C1r.
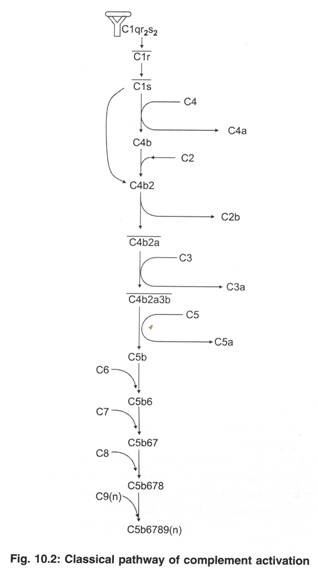
Cir memotong C1s. C1 yang dibelah menjadi enzim C1 yang dirancang aktif.
sebuah†”
C1 pada gilirannya membelah dua komponen komplemen, C4 dan C2.
saya. C4 dibelah menjadi fragmen C4a dan C4b. Fragmen C4b menempel pada permukaan sel mikroba.
- Komponen C2 melekat pada C4b. Komponen C2 yang melekat pada C4b dibelah menjadi fragmen C2a dan C2b oleh C1s. Fragmen C2b berdifusi meninggalkan kompleks C4b2a pada permukaan sel mikroba.
sebuah†”
Kompleks C4b2a bekerja pada C3 dan membelah C3 menjadi fragmen C3a dan C3b. (Karena kompleks C4b2a membelah C3, kompleks C4b2a juga disebut C3 convertase.)
sebuah†”
Fragmen C3b berikatan dengan C4b2a dan membentuk kompleks C4b2a3b. (Beberapa fragmen C3b mengikat permukaan sel target dan bertindak sebagai opsonin untuk fagositosis sel target.
sebuah†”
Kompleks C4b2a3b membelah C5 menjadi C5a dan C5b. (Karena C4b2a3b memotong C5, kompleks C4b2a3b disebut jalur klasik C5 convertase.) Fragmen C5b berikatan dengan permukaan mikroba.
sebuah†”
C6 berikatan dengan C5b dan membentuk kompleks C5b6.
sebuah†”
C7 berikatan dengan C5b6 dan membentuk kompleks C5b67. Daerah hidrofobik kompleks C5b67 berikatan dengan fosfolipid membran sel mikroba dan kompleks C5b67 dimasukkan ke dalam membran sel mikroba.
sebuah†”
C8 berikatan dengan C5b67 dan membentuk kompleks C5b678. Kompleks C5b678 menciptakan pori kecil (diameter 10-A) di membran sel mikroba.
sebuah†”
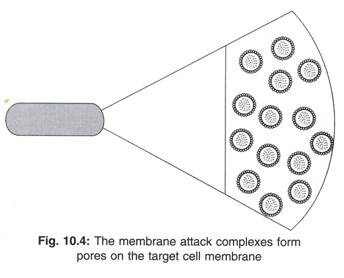
Banyak molekul (10 hingga 17 molekul) C9 berikatan dengan satu C5b678 untuk membentuk kompleks C5b6789 (n). Kompleks C5b6789 (n) juga disebut sebagai kompleks serangan membran (MAC). Kompleks MAC meningkatkan ukuran pori menjadi 70-100 A pada membran sel mikroba (Gambar 10.3). Banyak MAC terbentuk selama aktivasi komplemen dan setiap MAC mampu melubangi membran sel (Gambar 10.4). Karena tekanan osmotik yang tinggi di dalam sel mikroba, air dari luar masuk ke dalam mikroba. Akibatnya, sel mikroba membengkak dan pecah (yaitu mikroba lisis).
Gambar 10.3: Kompleks serangan membran .
Kompleks C5b6789 (n) yang dibentuk oleh aktivasi komplemen juga disebut kompleks serangan membran (MAC). MAC adalah produk seperti silinder dan melubangi membran sel. Melalui silinder seperti cairan lubang dan molekul mengalir masuk dan keluar dari sel, mengakibatkan kematian sel
Dengan demikian aktivasi jalur komplemen klasik oleh antibodi yang terikat antigen menghasilkan lisis sel mikroba yang mengekspresikan antigen. Karena jalur klasik diprakarsai oleh antibodi, jalur klasik berperan dalam respon imun yang didapat. Dengan tidak adanya antibodi spesifik terhadap mikroba (yang masuk ke dalam tubuh), jalur komplemen klasik tidak akan diaktifkan (meskipun semua komponen komplemen yang dibutuhkan ada di dalam tubuh).
Fragmen komplemen lainnya (seperti C4a, C3a, dan C5a) yang terbentuk selama aktivasi komplemen memiliki banyak fungsi penting dan akan dijelaskan kemudian (Tabel 10.2).
Jalur Alternatif Aktivasi Komplemen:
Berbeda dengan jalur komplemen klasik, jalur komplemen alternatif tidak memerlukan antibodi terhadap antigen untuk inisiasi aktivasi komplemen. Ini berimplikasi bahwa jalur alternatif diaktifkan bahkan selama masuknya antigen pertama kali. Dengan kata lain, jalur alternatif diaktifkan selama respon imun bawaan. Jalur komplemen alternatif memainkan peran pertahanan penting melawan mikroba segera setelah mikroba masuk ke inang.
C3, faktor B, faktor D, dan properdin adalah empat protein serum yang terlibat dalam inisiasi aktivasi jalur komplemen alternatif (Gambar 10.5; Tabel 10.3).
Molekul C3 memiliki ikatan thioester yang tidak stabil. Karena sifat ikatan tioester yang tidak stabil, C3 dalam darah secara spontan terhidrolisis menjadi C3a dan C3b. Jika mikroba kebetulan ada di dekat tempat pembentukan C3b, fragmen C3b menempel pada permukaan sel mikroba.
sebuah†”
Faktor B berikatan dengan C3b pada permukaan mikroba.
sebuah†”
Faktor D bekerja secara enzimatik pada faktor B yang terikat C3b untuk menghasilkan dua fragmen, fragmen Ba dan fragmen Bb. Fragmen Ba berdifusi dan kompleks C3bBb terbentuk. Kompleks C3bBb memiliki waktu paruh hanya 5 menit. Tetapi pengikatan protein serum lain yang disebut properdin memperpanjang waktu paruh C3bBb hingga 30 menit.
sebuah†”
Kompleks C3bBb memotong molekul C3 lain untuk menghasilkan fragmen C3a dan C3b. (Kompleks C3bBb disebut jalur alternatif C3 convertase.) Fragmen C3b berikatan dengan C3bBb dan membentuk kompleks C3bBb3b.
sebuah†”
Kompleks C3bBb3b membelah C5 menjadi C5a dan C5b (sehingga C3bBb3b disebut sebagai jalur alternatif C5 convertase). Langkah-langkah aktivasi komplemen selanjutnya mirip dengan langkah-langkah dalam jalur aktivasi komplemen klasik.

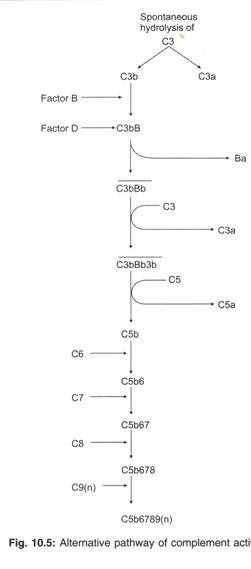
C6 berikatan dengan C5b dan membentuk kompleks C5b6.
sebuah†”
C7 berikatan dengan C5b6 dan membentuk kompleks C5b67.
sebuah†”
C8 berikatan dengan C5b67 dan membentuk kompleks C5b678.
sebuah†”
Banyak molekul C9 berikatan dengan C5b678 dan membentuk kompleks C5b6789 (n) (kompleks serangan membran). Kompleks serangan membran melubangi dinding sel mikroba dan menyebabkan lisis mikroba.

Amplifikasi Langkah Aktivasi Pelengkap:
Komponen komplemen dalam darah berada dalam keadaan tidak aktif secara fungsional. Beberapa komponen komplemen adalah proenzim. Ketika proenzim dibelah menjadi dua fragmen, salah satu fragmen memperoleh aktivitas enzimatik.
Setiap molekul enzim yang terbentuk pada setiap tahap aktivasi komplemen bekerja pada banyak molekul komponen komplemen berikutnya, menghasilkan aktivasi banyak komponen komplemen. Dengan demikian jumlah komponen komplemen yang diaktifkan pada setiap langkah meningkat berkali-kali lipat sehingga dihasilkan sejumlah besar kompleks penyerang membran dan fragmen komplemen lainnya, (misalnya satu molekul C3 convertase dapat bekerja pada 200 molekul C3 dan menghasilkan 200 fragmen C3b.) Amplifikasi semacam itu aktivasi komplemen pada berbagai langkah membantu inang dalam memproduksi pertahanan termediasi komplemen yang efektif.
Aktivasi Jalur Komplemen Klasik Non-imunologis:
Biasanya, aktivasi jalur komplemen klasik dimulai dengan pengikatan C1q ke antibodi yang sudah terikat pada antigen (yaitu inisiasi jalur klasik aktivasi komplemen berbasis imun). Namun, aktivasi jalur komplemen klasik juga dapat dimulai dengan cara non-imunologis.
saya. Beberapa bakteri (seperti Esch.coli dan beberapa strain Salmonella) dan virus (seperti virus Para influenza dan HIV) secara langsung berikatan dengan Clq dan memulai aktivasi komplemen. Aktivasi jalur klasik non-imunologi seperti itu mungkin berguna bagi inang a) untuk bertindak melawan mikroba sebagai respon imun bawaan, dan b) untuk bertindak melawan mikroba bahkan sebelum antibodi berikatan dengan mikroba.
- Kristal urat, endotoksin bakteri, dan heparin juga dapat menginisiasi aktivasi jalur komplemen klasik secara non-imunologis.
2. Jalur Aktivasi Pelengkap Lektin:
Baru-baru ini, jalur aktivasi komplemen ketiga yang disebut ‘jalur lektin aktivasi komplemen’ telah dijelaskan. Lektin adalah protein yang berikatan dengan karbohidrat. Langkah-langkah jalur lektin mirip dengan jalur klasik kecuali jalur lektin tidak memerlukan antibodi untuk memulai aktivasi komplemen.
Mannose binding lectin (MBL) adalah protein fase akut yang diproduksi selama respon inflamasi akut. MBL memiliki struktur yang mirip dengan C1q. Dua molekul lain yang disebut protease serin 1 dan 2 terkait MBL (MASP-1 dan MASP-2) dikaitkan dengan aktivasi sistem komplemen MBL. Protease serin terkait MBL dan MBL diyakini bertindak sebagai C1q, C1r, dan C1s dari jalur komplemen klasik.
MBL berikatan dengan molekul karbohidrat pada permukaan sel bakteri.
sebuah†”
Kemudian protease serin 1 dan 2 terkait MBL diaktifkan, yang mengarah ke pembelahan C4 dan C2.
sebuah†”
Langkah selanjutnya mirip dengan aktivasi komplemen klasik.
Namun, banyak detail jalur lektin yang belum diketahui. Karena jalur lektin tidak memerlukan molekul antibodi spesifik untuk aktivasi, disarankan bahwa jalur lektin aktivasi komplemen mungkin merupakan salah satu mekanisme pertahanan bawaan yang penting. MBL mengenali berbagai bakteri, virus, jamur, dan parasit yang signifikan secara klinis.
Mutasi dalam gen MBL diketahui menghasilkan kadar MBL plasma yang kurang optimal, atau defisiensi MBL. Karena tindakan yang tumpang tindih dari banyak mekanisme kekebalan lainnya, kadar MBL yang rendah biasanya tidak menyebabkan gejala klinis pada individu yang imunokompeten. Tetapi defisiensi MBL merupakan faktor risiko yang signifikan untuk infeksi pada pasien dengan gangguan kekebalan (seperti pasien kanker yang menjalani kemoterapi).