Beberapa jenis utama aberasi kromosom adalah sebagai berikut:
Susunan dan keberadaan banyak gen pada satu kromosom memberikan perubahan informasi genetik tidak hanya melalui perubahan jumlah kromosom tetapi juga melalui perubahan struktur kromosom.
Gambar milik: neurorexia.files.wordpress.com/2013/05/figure-1-histones-1024×1022.jpg
Perubahan kromosom disebabkan oleh perubahan materi genetik melalui kehilangan, penambahan atau penataan ulang segmen tertentu. Perubahan seperti itu disebut penyimpangan kromosom. Modifikasi membawa mutasi kromosom. Mutasi kromosom sangat jarang di alam tetapi dapat dibuat secara artifisial oleh sinar ‘X’, radiasi atom dan bahan kimia, dll.
Perubahan struktural pada kromosom disebabkan oleh pecahnya kromosom, atau pada subunit pembelahan selnya, yaitu kromatid. Setiap jeda menghasilkan 2 ujung yang kemudian dapat mengikuti tiga jalur berbeda. (Gbr.43.1).
(a) Mereka dapat bersatu kembali, yang pada akhirnya menyebabkan hilangnya segmen kromosom yang tidak mengandung sentromer.
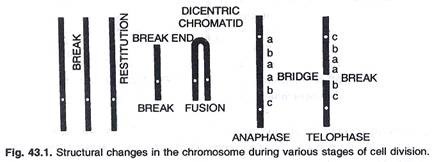
(b) Penyatuan kembali atau penyusunan kembali yang segera dari ujung-ujung yang rusak dapat terjadi, yang mengarah ke penyusunan kembali struktur aslinya.
(c) Satu atau kedua ujung dari satu pemutusan tertentu dapat bergabung dengan yang dihasilkan oleh pemutusan lain yang menyebabkan pertukaran, atau penyatuan non rekonstitusional.
Mc Clintock (1941) mempelajari di Zea Mays bahwa kromosom pecah dan mengikuti duplikasi. Sebuah kromatid disentrik ditemukan. Selama serat gelendong anafase melekat pada dua sentromer menghasilkan pembentukan jembatan dari satu kutub ke kutub lainnya. Jembatan putus menyebabkan kekurangan atau duplikasi.
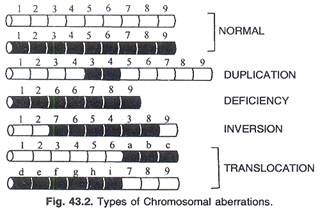
Penyimpangan kromosom terdiri dari 4 jenis utama:
(a) Penghapusan (b) duplikasi (c) inversi dan (d) translokasi. (Gbr. 43.2).
(A) Penghapusan atau Kekurangan:
Delesi atau defisiensi sesuai dengan namanya adalah hilangnya segmen kromosom. Setelah istirahat bagian tanpa sentromer hilang. Di sisi lain, bagian yang menempel pada sentromer bertindak sebagai kromosom yang kekurangan. Bridges (1917) untuk pertama kalinya mengamati defisiensi di lokus Bar dari Drosophila.
Dua jenis penghapusan ditemukan:
Penghapusan terminal:
Putus tunggal di dekat ujung kromosom. Dijelaskan dalam jagung tetapi sebaliknya tidak umum.
Penghapusan pengantara:
Kromosom pecah dan bersatu kembali tetapi bagian itu hilang di antara keduanya. (Gbr. 43.3). Penghapusan terdeteksi pada saat pasangan homolog. Jika ada bagian kromosom yang hilang maka kromosom yang lain juga harus menghilangkannya dalam bentuk tonjolan untuk membuat sinapsis. misalnya, jika kromosom memiliki 1, 2, 3, 4, gen. Bagian 2 hilang dari satu kromosom yang keluar, 1, 3, 4. Kromosom homolog lainnya pada saat sinaps menonjol keluar atau membentuk lingkaran pada posisi 2.
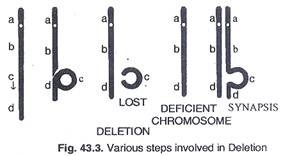
Jika segmen yang hilang memiliki kepentingan fisiologis, individu tersebut tidak akan bertahan. Jika gen dominan ‘A’ hilang, alel resesif ‘a’ dapat mengekspresikan dirinya. Ini disebut dominasi semu.
Pada manusia, penghapusan kromosom 5 mengakibatkan sindrom cri-du-chat, anak-anak menangis seperti kucing, berkepala kecil dan mengalami keterbelakangan mental.
Penghapusan sebagian kromosom ke-18 menghasilkan sindrom telinga besar dan jari panjang.
Pada jagung kekurangannya terbatas pada sterilitas polen. Gemetophyte haploid jantan menunjukkan defisiensi sedangkan betina mungkin menerima metabolit dari jaringan ibu yang melengkapi kekurangan tersebut. Segmen yang dihilangkan membentuk gesper. (Gbr. 43.4)
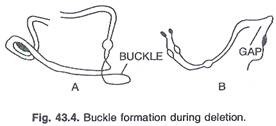
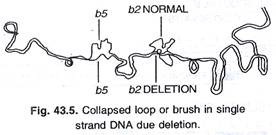
Kekurangan E. coli juga dicatat. Penghapusan menunjukkan bahwa DNA itu beruntai tunggal dan terlihat seperti lingkaran atau sikat yang runtuh. (Gbr. 43.5).
(B) Duplikasi:
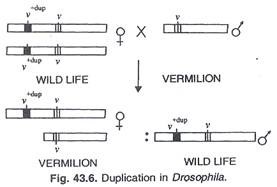
Berikut segmen kromosom diulang dua kali, yaitu, digandakan. Duplikasi ditemukan pada kromosom Drosophila ‘X’ untuk pertama kalinya yang membawa alel tipe liar untuk vermilion (v + ) dan telah dialihkan ke kromosom ‘X’ yang membawa alel vermilion mutan (v), Bridges menemukan bahwa karena fakta bahwa Kromosom ‘X’ membawa alel v dan v + keduanya wild type bukan vermilion. Properti yang sama dari v dan v + menghasilkan efek tipe liar. ‘Betina duplikasi’ seperti itu ketika disilangkan dengan laki-laki vermilion nonduplikasi semua keturunan perempuan adalah vermilion dan semua keturunan laki-laki, yaitu y adalah tipe liar. (Gbr.43.6.)
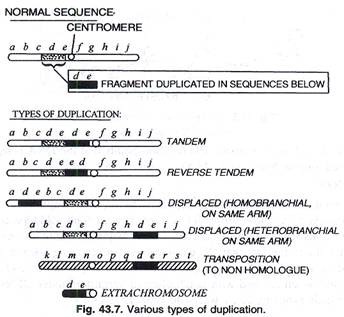
Jenis duplikasi:
Duplikasi terdiri dari berbagai jenis. (Gbr. 43.7)
Duplikasi tandem:
Ketika segmen duplikasi dekat sentromer misalnya, urutan pada kromosom isabcdefghithe sentromer hadir antara e dan f segmen de diulang segera setelah posisi normalnya.
Tandem terbalik:
Bila ruas dibalik dalam duplikasi, misalnya de ruas yang digandakan maka akan digandakan sebagai akta bukan ded e.
Tandem terlantar:
Segmen diulangi di suatu tempat yang jauh dari lokasi aslinya tetapi di lengan yang sama (perpindahan homobrakial) atau di lengan lainnya (perpindahan heterobrakial).
Transposisi:
Ketika segmen diduplikasi pada kromosom non homolog itu disebut transposisi.
Kromosom ekstra:
Duplikasi melibatkan sentromer yang disebut kromosom ekstra. Dalam duplikasi kromosom kelenjar ludah umum terjadi baik sebagai tekuk dalam heterozigot duplikasi atau sebagai pasangan silang antara bagian-bagian dari kromosom yang berbeda.
(C) Translokasi:
Transfer bagian dari satu kromosom ke kromosom non homolog dikenal sebagai translokasi. Ketika ada pertukaran segmen pada dua kromosom non homolog itu disebut translokasi resiprokal. Ini juga mencakup pertukaran segmen antara bagian non homolog dari sepasang kromosom, misalnya kromosom ‘X’ atau ‘Y’. Segmen tersebut tidak hilang atau ditambahkan, melainkan hanya ditukar.
Ini pertama kali diamati di Drosophila oleh perilaku yang tidak biasa dari gen kromosom ke-2 tertentu yang disebut Pucat. Itu mematikan dalam kondisi homozigot. Bridges mengamati bahwa sifat mematikannya dapat ditekan dengan adanya gen lain pada kromosom ke-3 yang juga mematikan dalam kondisi homozigot. Efek pucat disebabkan karena kekurangan ujung kecil kromosom ke-2 termasuk pleksus atau balon yang terhubung ke gen kromosom ke-3 antara kayu hitam atau kasar.

Stern pada tahun 1926 mengamati translokasi beberapa alel (bobbed) pada kromosom ‘Y’ ke kromosom ‘X’. (Gbr.43.8)
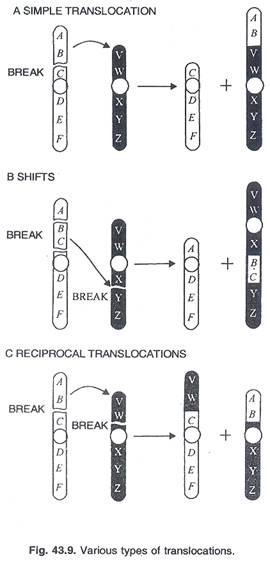
Jenis translokasi:
(a) Translokasi sederhana:
Pemutusan tunggal pada kromosom dan dipindahkan ke ujung yang lain. (Gbr. 43.9)
(b) Pergeseran atau translokasi interkalar:
Jenis translokasi umum yang melibatkan 3 patahan sehingga dua bagian patahan dari satu kromosom (misalnya Pale) disisipkan di dalam patahan yang dihasilkan dalam kromosom non homolog. (Gbr. 43.9B)
(c) Translokasi timbal balik atau Interchange:
Translokasi yang sering diamati di mana pemutusan tunggal dalam dua kromosom homolog menghasilkan pertukaran segmen kromosom di antara mereka. (Gbr. 43.9c)
Homozigot translokasi membentuk jumlah pasangan homolog yang sama dengan homozigot normal selama sentromer tidak hilang.
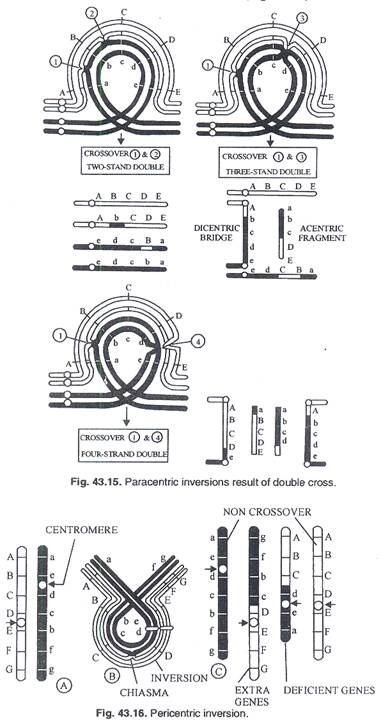
Hasil pasangan dan meiosis berbeda dalam translokasi heterozigot yang mengandung dua segmen translokasi dan pasangan normalnya. (Gbr. 43.10). Translokasi timbal balik membentuk kompleks 4 kromosom pada tahap pachytene. Chiasmata antara kromosom tersebut dapat membentuk kuadrivalen yang kemudian dapat dipisahkan dalam 3 pola segregasi yang berbeda pada pembelahan meiosis pertama. (Gbr. 43.11).
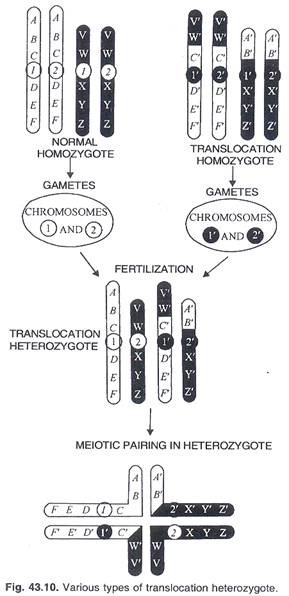
(i) Segregasi alternatif:
Sentromer nonhomolog berlawanan atau bergantian pergi ke kutub yang sama secara zigzag, sehingga kromosom nontranslokasi (1, 2) dan translokasi (1′, 2′) berada dalam gamet yang terpisah. Gamet memiliki komplemen gen yang seimbang lengkap tanpa duplikasi atau defisiensi (Gambar 43.11).
(ii) Segregasi yang berdekatan-1:
Kromosom berdekatan yang tidak homolog pergi ke kutub yang sama tetapi setiap gamet mengandung kromosom translokasi dan non translokasi (1 2′, 1’2) kedua defisiensi duplikasi pada setiap gamet ada (Gbr. 43.11).
(iii) Pemisahan yang berdekatan-2:
Sentromer yang berdekatan kembali ke kutub yang sama tetapi ini sekarang homolog serta mengandung kromosom translokasi dan non translokasi (1, 1′; 2, 2′). Duplikasi dan defisiensi menghasilkan komponen gen yang tidak seimbang (Gambar 43.11C).
Segregasi yang berdekatan-1 dan yang berdekatan-2 menghasilkan gamet yang tidak seimbang. Gamet subur dari heterozigot translokasi sebagian besar akan terbatas pada segregasi alternatif.
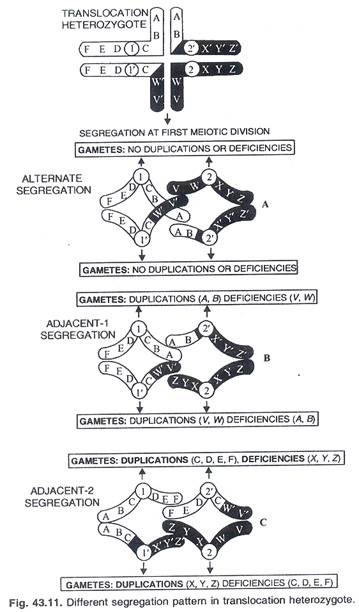
Sinapsis dari kromosom translokasi heterozigot yang menunjukkan konfigurasi seperti silang kemudian membuka sebuah cincin atau angka delapan (Gambar 43.12).
Konsekuensi dari segregasi tersebut adalah pemilahan independen antara gen dan kromosom nonhomolog akan terhambat. Karena duplikasi dan defisiensi, tak satu pun dari fenotipe mutan tunggal akan muncul pada keturunannya. Heterozigot translokasi memiliki kesuburan rendah. Jika tingkat duplikasi dan defisiensi kecil, gamet atau zigot yang tidak seimbang belum tentu mematikan.
(D) Pembalikan:
Bagian kromosom yang berubah akibat rotasi 180° disebut inversi. Urutan gen di dalamnya dibalik.
Inversi muncul dengan pembentukan loop pada kromosom. Terobosan dapat terjadi pada titik persimpangan loop (Gbr. 43.13). Penyatuan kembali ujung yang putus terjadi dalam kombinasi baru, dan terbalik. Heterozigot inversi dibentuk oleh loop dan tonjolan berpasangan.
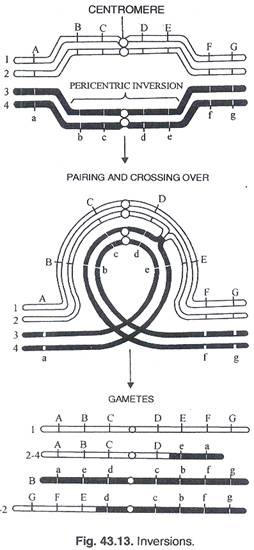
Jenis Inversi:
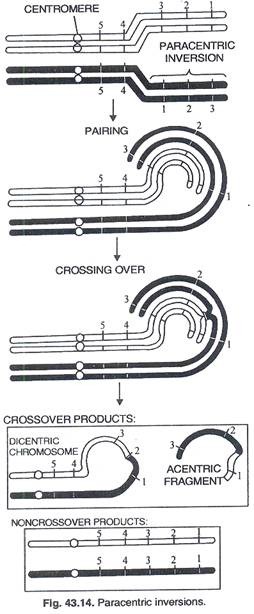
Inversi parasentrik: Pelaku segmen terbalik no; termasuk sentromer. Inversi perisentrik: Segmen terbalik termasuk sentromer.
Inversi parasentrik:
Persilangan tunggal pada daerah terbalik akan menghasilkan pembentukan kromosom disentrik (dengan 2 sentromer) dan kromosom asentrik (tanpa sentromer). Dari 2 kromatid yang tersisa, satu akan normal dan yang lainnya membawa inversi. Kromatid disentrik dan kromatid asentrik akan diamati pada anafase I berupa jembatan dan fragmen (Gambar 43.14). Crossover ganda menunjukkan defisiensi dan duplikasi (Gambar 43.15) sehingga menimbulkan variasi konfigurasi anafase I.
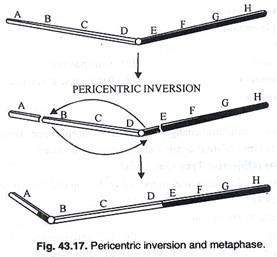
Inversi perisentrik:
Dalam sentromer inversi pericentric berada di segmen terbalik. Pada pakiten tahap 2 dari 4 kromatid yang dihasilkan setelah meiosis akan mengalami defisiensi dan duplikasi. Tidak ada jembatan disentrik atau fragmen asentrik yang akan diamati (Gbr.43.16). Dalam inversi perisentrik, jika dua patahan tidak terletak sama jauhnya dari sentromer, akan terjadi perubahan bentuk kromosom. Kromosom metasentrik dapat menjadi submetasentrik dan sebaliknya (Gambar 43.17).