Baca artikel ini untuk mempelajari tentang Komponen Sel Hidup dan Fungsinya!
Struktural Sel:
Sel adalah unit struktur dan aktivitas biologis. Ini terdiri dari massa protoplasma terorganisir yang dikelilingi oleh penutup pelindung dan permeabel selektif yang disebut membran plasma.
Protoplasma suatu sel disebut protoplas (Hanstein, 1880). Itu terbuat dari plasma-lemma, sitoplasma, vakuola dan nukleus. Pada sel tumbuhan, jamur, bakteri dan cyanobacteria, sel juga dikelilingi oleh dinding sel.

Sel adalah unit struktur dan aktivitas biologis. Ini terdiri dari massa protoplasma terorganisir yang dikelilingi oleh penutup pelindung dan permeabel selektif yang disebut membran plasma. Protoplasma suatu sel disebut protoplas (Hanstein, 1880). Itu terbuat dari plasma-lemma, sitoplasma, vakuola dan nukleus. Pada sel tumbuhan, jamur, bakteri dan cyanobacteria, sel juga dikelilingi oleh dinding sel.
[I] Protoplasma:
Protoplasma digambarkan sebagai ‘dasar fisik kehidupan’ oleh Thomas Huxley (1868), karena semua aktivitas makhluk hidup adalah aktivitas protoplasma itu sendiri. Ini adalah satu-satunya zat yang diberkahi dengan kehidupan dan, oleh karena itu tumbuhan dan hewan yang mengandung zat ini di dalam tubuhnya dianggap hidup. Saat protoplasma mati, sel berhenti melakukan fungsi apa pun untuk tumbuhan atau hewan, yang kemudian secara keseluruhan menjadi tidak aktif dan mati.

Sifat Fisik Protoplasma:
Berdasarkan studi mikroskopis, protoplasma tampak sebagai zat gelati nous yang bening, tidak berwarna, seperti jeli. Ini adalah larutan encer dari butiran yang sangat kecil dan butiran yang berbeda (zat karena ini tampak kental, elastis dan butiran).
Dalam keadaan aktif protoplasma tetap jenuh dengan air yang merupakan 75-90 persen kandungannya. Protoplasma menggumpal saat dipanaskan, dan ketika dimatikan, ia kehilangan transparansi. Menanggapi berbagai rangsangan eksternal, misalnya, sengatan listrik dan bahan kimia.
Protoplasma bersifat semi-permeabel dan sifat ini hilang ketika protoplasma dibunuh. Dalam kondisi normal, protoplasma sel hidup berada dalam keadaan bergerak lambat tetapi konstan. Namun, dalam banyak kasus, ini menunjukkan gerakan berbeda dari jenis yang berbeda.
Massa protoplasma telanjang, menunjukkan dua jenis gerakan — ciliary (gerakan berenang dari tubuh protoplasma yang bebas, kecil seperti zoospora dari banyak ganggang dan jamur dengan bantuan silia atau flagela) dan amoeboid (gerakan merayap dengan bantuan pseudopodia seperti pada Amoeba). Protoplasma, yang tertutup oleh dinding sel, menunjukkan gerakan mengalir di dalamnya, yang disebut sebagai siklosis.
Cyclosis terdiri dari dua jenis-rotasi (protoplasma bergerak di sekitar vakuola di dalam sel hanya dalam satu arah, baik searah jarum jam atau berlawanan arah jarum jam, misalnya, sel daun Hydrilla atau Vallisneria dan sirkulasi (protoplasma bergerak dalam arah yang berbeda di sekitar yang berbeda). vakuola di dalam sel seperti pada rambut staminal Tradescantia).
Protoplasma adalah sistem koloid polifasik yang kompleks. Karakteristik fisik yang paling penting dari protoplasma adalah sifat koloidnya yang menjadi ciri utamanya.
Komposisi Kimia Protoplasma:
Protoplasma aktif mengandung persentase air yang tinggi (75-90%) dan tetap jenuh dengannya. Meninggalkan air ini, materi padat protoplasma mengandung yang berikut: protein-40-60%.
Zat berlemak (lemak dan lipid sejati, terutama lesitin) -12-14%; karbohidrat-12-14% dan garam anorganik 5-7%. Analisis protoplasma mati mengungkap daftar panjang unsur-unsur yang ada di dalamnya.
Oksigen (O) sekitar 65%’, karbon (C) sekitar 18,3%, hidrogen (H) sekitar 11% dan nitrogen (N) sekitar 2,5%, yang paling mencolok. Unsur lain yang membentuk 3% sisanya termasuk klorin (CI), belerang (S), fosfor (P), silikon (Si), kalsium (Ca), mangan (Mn), dll.
Selain bahan kimia yang disebutkan di atas, berbagai jenis pigmen, lateks, vitamin, zat pengatur tumbuh, enzim dan alkaloid juga ditemukan dalam protoplasma dan memainkan peran khusus dalam sel.
Protoplasma memiliki dua bagian utama
- Sitoplasma
- Inti
Bagian terluar sitoplasma terdiri dari membran satuan yang disebut plasma-lemma atau membran plasma pada sel hewan dan tumbuhan. Pada sel tumbuhan, membran plasma dilindungi dari luar oleh dinding sel yang tebal.
[II] Dinding sel:
Dinding sel adalah karakteristik sistem tidak hidup dari sel tumbuhan saja. Secara luar biasa, gamet tumbuhan rendah tertentu dan plasmodium jamur tertentu tidak memiliki dinding sel. Hanya ketika dinding sel bersentuhan dengan protoplas hidup yang membentuknya, ia mampu tumbuh sehingga tidak dapat dikatakan tidak bergantung pada protoplas.
Dinding Sel Terdiri Dari Tiga Bagian:
1. Lamela tengah:
Ini tipis, amorf, membentuk zat antar sel dan membantu menyatukan dinding primer sel yang berdekatan. Lamela tengah tidak ada di sisi luar sel permukaan. Itu terdiri dari kalsium dan magnesium pektat. Lamela tengah dapat dilarutkan oleh berbagai zat termasuk pektinase dan cairan Jeffery. Secara optik isotropik.
2. Dinding utama:
Ini adalah yang pertama dibentuk oleh sel dan disimpan di kedua sisi lamela tengah dalam sel yang berdekatan. Ini secara optik anisotropik dan secara kimiawi terdiri dari selulosa, zat pektik, hemiselulosa dan beberapa polisakarida lainnya. Semua sel hidup memiliki dinding primer. Karena dinding primer diendapkan sangat awal dalam ontogeni sel, ia mengalami tekanan yang cukup besar selama pertumbuhan sel selanjutnya.
Itu tidak hanya tumbuh dalam dimensi linier tetapi juga dalam ketebalan. Dengan demikian, dinding sel primer memiliki plastisitas dan elastisitas. Dalam beberapa kasus, pertumbuhan ketebalan dinding primer bersifat reversibel berbeda dengan sifat dinding sekunder yang ireversibel.

Dinding primer terdiri dari sejumlah mikrofibril yang tertanam dalam matriks seperti gel amorf atau zat dasar. Di sebagian besar tanaman, mikro-fibril terbentuk dari selulosa. Mereka 1 terbuat dari polimer asetil glukosamin atau selulosa jamur di banyak jamur. Mikro-fibril diorientasikan secara bervariasi sesuai dengan bentuk dan penebalan dinding. Biasanya mereka diatur dalam jaringan yang longgar.
3. Dinding Sekunder:
Ini diproduksi di beberapa sel dewasa ketika kemudian berhenti tumbuh, misalnya trakeid, elemen pembuluh, serat, kolenkim. Dinding sekunder diletakkan di bagian dalam dinding primer dengan akresi atau pengendapan material di atas permukaan struktur yang ada. Tebal (3-10 μm) dan terdiri dari setidaknya tiga lapisan, kadang-kadang lebih (misalnya, tabung lateks dari Euphorbia mili).
Mereka diberi nama sebagai S 1 , S 2 , S 3 , S x dll. Lapisan paling dalam dari dinding sekunder kadang-kadang berbeda baik secara kimia maupun sifat pewarnaan karena adanya xilan. Ini kemudian disebut dinding tersier, misalnya kayu tegang pada gymnospermae. Dinding sekunder mungkin tidak ada, terendapkan secara tidak teratur atau terbentuk secara seragam di dalam sel. Ini menghasilkan diferensiasi sel-parenkim, kolenkim, sklerenkim, trakeid, dan pembuluh darah.
Komposisi dinding sekunder pada dasarnya mirip dengan dinding primer karena memiliki mikrofibril selulosa yang tertanam dalam matriks pektin dan hemiselulosa. Mikro-fibril selulosa dari dinding sekunder terletak dekat, sejajar dan pada sudut sumbu longitudinal sel.
Sejumlah bahan yang berbeda dapat disimpan di dinding seperti lignin, suberin dan kutin. Zat lain yang dapat disimpan di dinding sel adalah silika (misalnya rumput), mineral, lilin, tanin, resin, gom, dll.
Komposisi Kimia Dinding Sel:
Dinding sel tanaman selain jamur terdiri dari selulosa-polisakarida. Rumusnya adalah (C 6 H J0 O 5 ) n. Selain selulosa, beberapa zat pectic, hemiselulosa dan karida polisakarida lainnya juga dapat hadir. Lamela tengah terbuat dari kalsium pektat dan magnesium pektat. Dinding primer dan sekunder biasanya terdiri dari selulosa meskipun beberapa zat lain mungkin juga ada.
Susunan Molekul dan Makromolekul Selulosa di dinding Sel:
Selulosa, polimer rantai lurus panjang d-glukosa, membentuk bagian yang baik dari kerangka dinding sel. Kira-kira 100 rantai panjang cellu hilang membentuk fibril elementer atau misel. Sekitar 20 fibril elementer merupakan mikro-fibril.
Ruang antara mikro-fibril diisi dengan air, zat pectic, hemiselulosa di dinding primer dan lignin, cutin, suberin, dll, di dinding sekunder. Sekitar 250 mi cro-fibril membuat fibril.

Plamodemata:
Plasmodesmata (tunggal— plasmodesma; Tangl, 1879; Strasburger, 1901) adalah jembatan sitoplasma antara sel tumbuhan yang berdekatan yang berkembang di pori-pori kecil dindingnya. Mereka membentuk kontinum protoplasma yang disebut symplast.
Berbagai zat dapat berpindah dari satu sel ke sel lainnya melalui plasmodesmata. Plasmodesma terdiri dari saluran yang dilapisi oleh membran plasma dan memiliki tubulus sederhana atau bercabang yang dikenal sebagai desmotubulus. Desmotubule adalah perpanjangan dari retikulum endoplasma.
Lubang:
Pit adalah area yang tidak menebal di dinding sekunder sel tumbuhan. Oleh karena itu, mereka muncul sebagai depresi. Pit umumnya terjadi berpasangan pada dinding dua sel yang berdekatan. Lubang memiliki rongga atau ruang lubang dan membran lubang. Membran pit terdiri dari dinding primer dan lamela tengah.
Lubang terdiri dari dua jenis, sederhana dan berbatasan. Lubang sederhana memiliki lebar ruang lubang yang seragam. Di lubang yang dibatasi, ruang lubang berbentuk labu karena dinding sekunder menutupi mulutnya.
Pit membran permeabel. Ini mungkin memiliki pori-pori submikroskopik kecil. Oleh karena itu, lubang membantu dalam translokasi cepat antara dua sel yang berdekatan.

Fungsi Dinding Sel:
Ia melakukan sejumlah fungsi:
(i) Melindungi protoplasma dari cedera mekanis.
(ii) Melindungi sel dari serangan patogen.
(iii) Memberikan kekakuan pada sel.
(iv) Memberikan bentuk pada sel dan melawan tekanan osmotik
(v) Memberi kekuatan pada tumbuhan darat untuk menahan gaya gravitasi.
(vi) Dengan pertumbuhannya, dinding membantu perluasan sel.
(vii) Tergantung pada jenis selnya, dindingnya mungkin permeabel atau impermeabel. Bahkan dinding permeabel memberikan beberapa efek pengaturan pada pengangkutan zat masuk dan keluar sel.
(viii) Lubang yang ada di dinding membantu menghasilkan kontinum atau simplas protoplasma di antara sel.
(ix) Cutin dan suberin dari dinding sel mengurangi kehilangan air melalui transpirasi.
(x) Dinding tabung ayakan, trakeid, dan bejana dikhususkan untuk transportasi jarak jauh.
[III] Sitoplasma:
Sitoplasma adalah massa protoplasma umum semi-cair seperti jeli tidak termasuk nukleus tetapi termasuk semua komponen lainnya – matriks sitoplasma, organel sel, dan inklusi sel.
- Matriks Sitoplasma atau Sitosol (Hyaloplasma):
Ini adalah bagian cairan bening dari sitoplasma yang dapat ada dalam dua keadaan, sol dan gel. Keduanya masing-masing disebut plasmasol dan plasmagel. Plasmagel biasanya hadir di bawah membran plasma. Ini disebut ektoplast.
Plasmasol internal dan dikenal sebagai endoplast. Air merupakan 90% dari matriks. Matriks sebenarnya adalah kompleks kristal-koloid dalam air di mana beberapa bahan kimia hadir dalam bentuk larutan sejati sementara yang lain hadir sebagai larutan koloid.
Matriks sitoplasma melakukan sejumlah fungsi. Yang penting adalah:
- Bahan pertukaran organel sel melalui matriks sitoplasma.
- Matriks mengandung bahan mentah dan menyediakan organel sel yang sama untuk fungsinya.
- Matriks adalah tempat sintesis sejumlah biokimia seperti lemak, nukleotida, beberapa karbohidrat, protein, koenzim, dll.
- Glikolisis, respirasi anaerobik dan jalur pentosa Jenis respirasi aerobik terjadi di bagian matriks sitoplasma.
- Membantu distribusi berbagai bahan di dalam sel.
- Organel Sel:
Mereka adalah struktur sub-seluler dengan bentuk morfologis yang khas, susunan kimiawi yang khas, dan fungsi yang pasti. Sebuah sel mengandung sejumlah organel seperti mitokondria, plastida, retikulum endoplasma, kompleks Golgi, ribosom, badan mikro, lisosom, dll.
- Mitokondria (Mito= benang, butiran kondrion)—Pertama kali ditemukan oleh Kolliker pada tahun 1880 dan digambarkan sebagai ‘bioplast’ oleh Altmann pada tahun 1894, organel ini diberi nama ulang mitokondria oleh Benda (1897).
Secara umum, mitokondria berbentuk batang tetapi sperma dan telur dari sejumlah organisme memiliki bentuk yang lebih bulat. Mitokondria memiliki kerangka lipoprotein yang mengandung banyak enzim dan koenzim yang diperlukan untuk metabolisme energi. Mereka juga mengandung DNA spesifik untuk pewarisan sitoplasma dan ribosom untuk sintesis protein.
Bentuk, ukuran, serta jumlahnya bervariasi tergantung jaringan, organisme, dan kondisi fisiologis organisme. Aktivitas sel adalah kriteria utama mitokondria untuk didistribusikan dalam sel misalnya, dalam sel hati mereka membentuk sekitar 30-35% protein sel sementara di sel ginjal mereka membentuk sekitar 20%.
Pada sel tumbuhan jumlahnya lebih sedikit dibandingkan dengan sel hewan. Meskipun didistribusikan secara merata di sitoplasma, mereka mungkin terbatas pada area tertentu di mana permintaan ATP yang tinggi harus dipenuhi, misalnya di sel otot, spermatozoa, batang dan kerucut mata. Mitokondria tidak ada pada prokariota dan eukariota anaerob.

Ultrastruktur Mitokondria:
Mitokondria tunggal memiliki diameter bervariasi antara 0,5 dan 1,0 μm dan panjang 7 μm. Drion mitokondria diapit oleh dua membran berbeda yang disebut membran luar dan dalam; yang terakhir memisahkan matriks organel, cairan seperti gel.
Dengan demikian membran dalam memisahkan matriks dari ruang membran dalam yang berisi cairan. Membran luar dan dalam mengandung berbagai enzim. Sejumlah enzim yang digunakan selama siklus Krebs melekat pada matriks dengan untaian DNA tersuspensi ke dalamnya dan ribosom tersebar di mana-mana.
Membran dalam dan luar sangat berbeda secara kimiawi dan struktural satu sama lain dan juga dalam permeabilitas. Membran bagian dalam memiliki luas permukaan yang lebih besar karena perluasannya ke dalam matriks dalam bentuk “krista” yang berbeda dalam bentuk dan jumlah, tergantung pada topografi seluler, mulai dari berbaring sejajar satu sama lain hingga membentuk jaringan anastomosis.
Jumlah krista bertambah atau berkurang tergantung pada aktivitas aerobik mitokondria karena selama aktivitas aerobik lebih banyak jumlah ATP yang akan diproduksi sehingga jumlah krista yang lebih besar untuk mengakomodasi sistem enzim. Membran bagian dalam kaya akan protein dan kardiolipin sedangkan bagian luarnya kaya akan fosfoloid. Kedua membran tampak pada pola trilamellar, dijelaskan oleh Robertson (1967).
Membran dalam dan juga krista-kristanya memiliki partikel kecil seperti raket tenis yang disebut partikel elementer, partikel F 0 – F 1 atau oksisom (=oksisom). Sebuah mitokondria mengandung partikel elementer lx 10 4 — lx 10 5 . Setiap partikel elementer atau oksisom memiliki kepala, tangkai, dan alas. Basis (subunit F 0 ) memiliki panjang sekitar 11 nm dan ketebalan 1,5 nm.
Tangkai panjangnya 5 nm dan lebar 3,5 nm. Kepala (subunit F 1 ) memiliki diameter 9—10 nm. Partikel elementer mengandung ATP-ase. Oleh karena itu, mereka adalah pusat sintesis ATP selama fosforilasi oksidatif. Enzim transpor elektron terletak di membran dalam yang bersentuhan dengan partikel elementer.
Di beberapa tempat, membran mitokondria luar dan dalam bersentuhan. Mereka disebut situs adhesi. Situs adhesi adalah daerah perembesan khusus mitokondria untuk transfer bahan dari luar ke dalam dan sebaliknya.
Ruang luar atau ruang peri-mitokondria adalah ruang yang terletak di antara membran luar dan dalam dari selubung mitokondria. Biasanya lebarnya 60-100 A° dan meluas ke ruang-ruang krista. Kamar berisi cairan yang memiliki beberapa enzim.
Ruang dalam membentuk inti mitokondria dan berisi matriks semi-cair. Matriks memiliki partikel protein, ribosom, RNA, DNA mitokondria, enzim siklus Krebs, sintesis asam amino dan metabolisme asam lemak. Ribosom mitokondria adalah 55 S hingga 70 S di alam. DNA telanjang. Ini umumnya melingkar tetapi bisa linier. DNA membuat mitokondria semi-otonom.
Status semi-otonom Mitokondria:
Mitokondria diketahui mengandung DNA dan RNA dan ada bukti yang menunjukkan bahwa mereka memiliki sintesis proteinnya sendiri. Namun, DNA inti juga berperan dalam sintesis protein mitokondria.
Untuk alasan ini mitokondria dianggap sebagai partikel otonom yang menyebar sendiri. Dipercayai bahwa protein struktural mitokondria dirakit in situ sementara protein larut dirakit pada ribosom non-mitokondria. Fungsi:
- Mitokondria adalah pabrik biokimia mini di mana bahan makanan atau substrat pernapasan dioksidasi sempurna menjadi karbon dioksida dan air. Energi yang dibebaskan dalam proses awalnya disimpan dalam bentuk koenzim tereduksi dan gugus prostetik tereduksi.
Yang terakhir segera mengalami oksidasi dan membentuk ATP yang kaya energi. ATP keluar dari mitokondria dan membantu melakukan berbagai proses yang membutuhkan energi sel seperti kontraksi otot, konduksi impuls saraf, biosintesis, transpor membran, pembelahan sel, gerakan, dll. Karena pembentukan ATP, mitokondria disebut pembangkit listrik sel.
- Mitokondria menyediakan perantara penting untuk sintesis beberapa bio-kimia seperti klorofil, sitokrom, pirimidin, steroid, alkaloid, dll.
- Matriks atau ruang dalam mitokondria memiliki enzim untuk sintesis asam lemak. Enzim yang dibutuhkan untuk pemanjangan asam lemak telah dilaporkan di ruang mitokondria luar.
- Sintesis banyak asam amino terjadi di mitokondria. Asam amino yang pertama kali terbentuk adalah asam glutamat dan asam aspartat. Mereka disintesis dari asam ∞-ketoglutarat dan asam oksaloasetat. Asam amino lainnya dihasilkan melalui transformasi dan trans samination atau transfer gugus amino (—NH 2 ) dari asam glutamat dan asam aspartat.
- Suatu organisme umumnya menerima mitokondria dari induknya, karena terdapat dalam telur.
- Plastida:
Istilah plastida diperkenalkan oleh E. Haeckel pada tahun 1866. Plastida adalah organel semi otonom yang memiliki DNA dan selubung membran ganda yang menyimpan atau mensintesis berbagai jenis senyawa organik. Dengan pengecualian beberapa protista, plastida hanya terbatas pada tanaman. Plastida berkembang dari prekursor tidak berwarna yang disebut proplastida.
Proplastida memiliki kemampuan untuk membelah dan berdiferensiasi menjadi berbagai jenis plastida. Tergantung pada warnanya, mereka terdiri dari tiga jenis utama – leukoplas, kromoplas, dan kloroplas (Schimper, 1883).
(i) Leukoplas (Gk. leucos—putih, plastos—dibentuk):
Mereka adalah plastida tidak berwarna yang umumnya terjadi di dekat nukleus dalam sel non-hijau dan memiliki lamellae internal. Grana dan pigmen fotosintesis tidak ada. Leukoplas memiliki ukuran dan bentuk yang bervariasi, misalnya bulat, lonjong, silindris, berserabut, dll.
Ada tiga jenis leukoplas khusus. (a) Amiloplas. Mereka adalah pati yang mengandung leukoplas. Ini mengandung butiran pati sederhana atau majemuk yang ditutupi oleh selubung protein khusus, misalnya Umbi kentang, Beras, Gandum, (b) Elaioplasts (Lipidoplasts, Oleoplasts). Plastida tidak berwarna menyimpan lemak, misalnya Tube Rose, (c) Aleuroplasts, Proteoplasts atau Proteinoplasts. Plastida mengandung protein dalam keadaan amorf, kristaloid atau kristalo-globoid (misalnya, sel aleuron dari biji Jagung, sel endosperma dari Castor).
(ii) Kromoplas (Gk. chroma—warna, plastos—dibentuk):
Plastida berwarna kuning atau kemerahan karena adanya pigmen karotenoid. Klorofil tidak ada. Chro moplasts terbentuk baik dari leucoplasts atau kloroplas. Perubahan warna dari hijau menjadi kemerahan selama pemasakan tomat dan cabai disebabkan oleh transformasi kloroplas menjadi kromoplas. Warna oranye akar wortel disebabkan oleh kromoplas.
(iii) Kloroplas (Gk. chloros = rumput hijau, cetakan plastos):
Mereka adalah plastida hijau yang memiliki pigmen fotosintesis, klorofil dan karotenoid, dan mengambil bagian dalam sintesis makanan dari bahan mentah anorganik dengan adanya energi radiasi. Kloroplas ganggang selain yang hijau disebut kromatofor (misalnya rhodoplast ganggang merah, phaeoplast ganggang coklat).
Di mana kloroplas hadir, jumlah, bentuk, dan ukurannya bervariasi. Pada tanaman yang lebih rendah, jumlahnya terlalu banyak per sel dan memiliki bentuk yang berbeda: berbentuk cangkir di Chlamydomonas, berbentuk korset di Ulothrix, berbentuk pita di Spirogyra, dll.
Dalam ganggang hijau, kloroplas dikaitkan dengan makanan dan menyimpan tubuh protein, yang disebut pirenoid. Tetapi pada sebagian besar tumbuhan hijau, ukurannya kecil, bikonveks, bertubuh diskrit, biasanya berkisar antara 4 hingga 6 μm.
Ultrastruktur Kloroplas:
Kloroplas tanaman hijau memiliki lapisan membran yang mirip dengan mitokondria, masing-masing setebal 50A dan ruang antar membran selebar 70-100A. Membran pembatas luar yang memberikan bentuk yang pasti pada kloroplas juga membantu mengatur pergerakan bahan, antara kloroplas dan sitoplasma di sekitarnya.
Membran dalam sejajar dengan membran luar dan dilemparkan ke dalam lipatan yang berjalan sejajar satu sama lain dan disebut lamellae. Sebagian besar lamellae dalam kloroplas tumbuhan tingkat tinggi tersusun dalam kantung berbentuk cakram yang disebut tilakoid. Thylakoids adalah elemen struktural kloroplas saat mereka mengambil bagian dalam fotosintesis.
Satu tumpukan tilakoid disebut granum. 40-100 grana dapat terjadi dalam kloroplas. Setiap granum memiliki 20-50 tilakoid. Grana tidak ada dalam kloroplas alga. Oleh karena itu, yang terakhir adalah agranal. Selaput Grana mengandung partikel protein dengan dua ukuran dasar, diameter 105 dan 140μ.
Partikel yang lebih besar diasosiasikan dengan fotosistem I fotosintesis dan yang lebih kecil diasosiasikan dengan fotosistem II fotosintesis. Karena adanya grana, tilakoid dibedakan menjadi tilakoid dua granal dan stroma atau tilakoid intergranal.
Stroma granular terdiri dari butiran pati dan pusat stroma; untaian DNA dan ribosom (70 S) juga tersebar melalui stroma. Kloroplas atau ct DNA telanjang, melingkar atau terkadang linier. Matriks chlo roplast tumbuhan tingkat tinggi dapat menyimpan pati sementara, sebagai butiran pati, yang dikenal sebagai pati asimilasi.

Membran grana lamellae terdiri dari protein dan fosfolipid dan mengandung klorofil dan karotenoid yang merupakan pigmen fotosintesis dasar.
Perkembangan Kloroplas:
Kloroplas baru dihasilkan oleh pembelahan kloroplas dewasa atau mereka berkembang secara de novo dari proplastida di dalam sel. Proplastida juga dapat menimbulkan jenis plastida lain selain kloroplas. Informasi genetik yang ada dalam inti sel dan kloroplas berinteraksi selama diferensiasi organel baru.
DNA yang terkandung dalam kloroplas itu sendiri ditranskripsi dan diterjemahkan untuk membentuk beberapa protein kloroplas dan sisa protein kloroplas diperoleh DNA inti setelah diterjemahkan.
Fungsi Kloroplas:
- Pada tumbuhan eukariotik, fotosintesis terjadi di kloroplas dengan adanya sinar matahari. Reaksi fotokimia (reaksi terang) fotosintesis terjadi pada membran tilakoid grana, sedangkan reaksi biokimia (reaksi gelap) terjadi di stroma.
- Kloroplas adalah mesin alami untuk sintesis senyawa organik kaya energi yang menyediakan makanan tidak hanya untuk tanaman tetapi juga untuk semua makhluk hidup.
- Selama proses fotosintesis kloroplas menyerap karbon dioksida (CO 2 ) dari lingkungan dan melepaskan oksigen (O 2 ) yang digunakan dalam respirasi sel oleh semua makhluk hidup. Dengan demikian ia menjaga keseimbangan CO 2 dan O 2 di lingkungan.
- Retikulum Endoplasma (ER):
Itu ditemukan secara independen oleh Porter (1945) dan Thompson (1945). Nama itu diberikan oleh Porter pada tahun 1953. Retikulum endoplasma adalah sistem 3-dimensi, rumit dan saling berhubungan dari saluran-saluran berselaput yang berjalan melalui sitoplasma. Di beberapa tempat, itu terhubung dengan plasma-lemma serta selubung nuklir. ER sel otot yang sangat berkembang disebut ‘retikulum sarkoplasma’.

Retikulum endoplasma dibedakan menjadi:
(a) Retikulum endoplasma kasar atau granular; kekasaran disebabkan oleh adanya partikel ribosom atau RNP (Ribonucleoprotein) dan
(b) Retikulum endoplasma halus atau agranular; itu halus karena tidak adanya ribosom.
Retikulum endoplasma kasar berkembang dengan baik dalam sel yang terlibat terutama dalam sintesis protein seperti sel penghasil enzim. Ini juga membantu menghasilkan lisosom. Membran mereka mengandung pori halus di area ribosom yang menempel untuk melewatkan polipeptida yang disintesis ke saluran retikulum endoplasma untuk transportasi.
Retikulum endoplasma kasar mengandung dua jenis glikoprotein (ribophorin I dan ribophorin II) untuk menempel pada ribosom. Retikulum endoplasma kasar sebagian besar terbuat dari cisternae. Tubulus sangat sedikit.
Retikulum endoplasma halus ditemukan berlimpah dalam sel yang mensintesis lipid dan pada sel yang memproduksi hormon steroid seperti kolesterol, gliserida, testosteron dan progesteron atau pada sel yang terlibat dalam metabolisme vitamin seperti sel epitel berpigmen retina. Itu sebagian besar terbuat dari vesikel dan tubulus. Sphaerosom diyakini berasal dari SER.
ER, dapat bertindak sebagai semacam sistem konduksi dan kontinuitas yang diamati dalam beberapa kasus antara ER dan selubung nukleus menunjukkan bahwa sifat ini mungkin dan ini mungkin salah satu mode untuk mengangkut RNA dan nukleo-protein dari nukleus ke nukleus. sitoplasma.
Struktur:
Retikulum endoplasma terdiri dari saluran atau ruang yang dilapisi membran yang berisi cairan yang disebut matriks endoplasma. Unit membran ER mirip dengan membran plasma dalam komposisi kimia dan tebal 50-60A 0 . Retikulum endoplasma dapat eksis dalam tiga bentuk— cisternae, vesikel dan tubulus. Ketiga bentuk ER dapat terjadi pada sel yang sama tetapi pada waktu perkembangan sel yang berbeda.
(i) Sisterna:
Mereka adalah bagian seperti kantung datar yang saling berhubungan dari retikulum endoplasma yang berdiameter 40-50 nm. Cisternae ditemukan dalam bundel di mana mereka terletak sejajar satu sama lain. Mereka terjadi di sel-sel yang secara aktif terlibat dalam aktivitas sintetik.

(ii) Vesikel:
Mereka adalah kantung oval atau bulat dengan diameter 25-500 nm. Vesikel muncul sebagai vakuola kecil. Mereka tetap terisolasi di sitoplasma. Vesikel disebut juga mikrosom.
(iii) Tubulus:
Mereka adalah ekstensi seperti tabung yang dapat dihubungkan dengan cisternae atau vesikel untuk membentuk sistem retikuler. Tubulus bisa tidak beraturan atau teratur, bercabang atau tidak bercabang dengan diameter 50-100 nm.
Fungsi:
Retikulum endoplasma sangat membantu dalam meningkatkan luas permukaan untuk aktivitas metabolisme yang memungkinkan enzim untuk bertindak dengan substratnya ke tingkat yang lebih besar.
Ini berfungsi sebagai kerangka kerangka sitoskeleton atau intraseluler dan ultra-struktural dengan memberikan dukungan mekanis untuk matriks sitoplasma koloid.
Rongga besar yang dihasilkan oleh penonjolan ER menyediakan tempat pengumpulan produk, sebelum diangkut keluar sel. Ini membuat konservasi produk metabolisme.
Membran retikulum endoplasma mengandung sejumlah enzim (misalnya ATP-ase, reduktase, dehidrogenase, fosfatase) untuk berbagai aktivitas metabolisme dan sitokrom yang berperan dalam transpor elektron.
- Kompleks Golgi:
Kompleks Golgi adalah struktur sitoplasma kompleks yang terdiri dari kantung membran halus atau cisternae, jaringan tubulus dengan vesikel dan vakuola, yang berperan dalam pembentukan, sekresi, dan produksi biokimia kompleks membran.
Itu dikelilingi oleh sitoplasma bebas organel yang disebut zona eksklusi atau zat dasar Golgi. Ini pertama kali dilihat oleh George (1867) tetapi dinamai ilmuwan Italia Camillo Golgi, yang pada tahun 1898 mengenali peralatan di sel saraf burung hantu dan kucing.
Aparatus atau kompleks Golgi tidak ada dalam sel prokariotik (PPLO, bakteri dan ganggang biru-hijau). Dalam sel hewan itu tunggal atau terdiri dari satu kompleks yang terhubung. Kedua kondisi tersebut masing-masing disebut terlokalisasi (sebagian besar sel vertebrata) dan menyebar (sebagian besar sel invertebrata, hati, dan sel saraf vertebrata). Pada sel tumbuhan terbentuk dari sejumlah unit yang tidak berhubungan yang disebut diktiosom.
Struktur:
Bentuk dan ukuran kompleks Golgi tidak tetap. Mereka bergantung pada keadaan fisiologis sel. Biasanya kompleks Golgi terdiri dari empat bagian—cisternae, tu bules, vesikel, dan vakuola.

Pinggiran cisternae melengkung lembut dan ini membuat seluruh tubuh Golgi tampak melengkung atau seperti busur; dengan demikian muka cembung cisternae memiliki “muka cis” atau “muka formasi” dan sisi cekung terdiri dari “muka trans” atau “muka maturitas”.
Sisterna mirip dengan retikulum endoplasma halus dalam penampilan dan membungkus lumen 60-90 A° yang mengandung zat cair atau matriks. Posisi wajah trans badan Golgi diatur secara khusus tergantung pada jenis selnya.
Karena hubungan morfologis dan fisiologisnya yang erat dengan retikulum endoplasma dan lisosom, ini kadang-kadang disebut sebagai elemen sistem endomembran atau vakuolar.
Kadang-kadang tepi cisternae berlubang, membentuk “fenestrae”. Jumlah dan ukuran badan Golgi bervariasi dengan aktivitas metabolisme sel. Selama pembelahan sel, jumlah badan Golgi meningkat karena ini mengeluarkan bahan untuk membentuk pelat sel.
Fungsi:
- Sejumlah protein yang disintesis oleh retikulum endoplasma kasar diteruskan ke badan Golgi untuk pemrosesan akhir sebelum diekspor ke tujuan akhir. Protein yang ditujukan untuk sekresi, perbaikan membran plasma, atau pengendapannya, mencapai permukaan cis aparatus Golgi.
Ketika berada di cisternae, protein ini difosforilasi, diglikolyasi, disulfasi, atau diproses secara sederhana. Glikolisis terjadi di wajah Trans dan di sinilah protein dipilah sesuai dengan tujuannya.
Fungsi dan efisiensi aparatus Golgi disebabkan oleh kelompok enzim yang disebut Nukleosida Difosfatase yang menghidrolisis difosfat uridin, guanosin, dan inosin.
- Pada oosit hewan, aparatus Golgi berfungsi sebagai pusat di mana kuning telur diendapkan, prosesnya disebut vitellogenesis.
- Sebagian besar karbohidrat kompleks, selain glikogen dan pati, disintesis di dalam kompleks Golgi, misalnya mukopolisakarida, asam hialuronat, hemiselulosa, dll.
Lisosom:
Mereka ditemukan secara tidak sengaja oleh Christian de Duve pada tahun 1955. Organel diamati di bawah mikroskop elektron oleh Novikoff (1956). Dia juga menciptakan istilah, Lisosom (Yunani lisis-pencernaan atau longgar, soma-tubuh) adalah vesikel kecil yang dibatasi oleh membran tunggal dan mengandung enzim hidrolitik dalam bentuk butiran kristal kecil atau butiran semi-kristal 5-8 nm. .
Membran berlapis tunggal terdiri dari lipoprotein dan sulit untuk mengidentifikasi lisosom berdasarkan karakter morfologi. Sekitar 40 enzim telah dicatat terjadi di dalamnya. Semua enzim tidak terjadi dalam lisosom yang sama tetapi ada kumpulan enzim yang berbeda dalam berbagai jenis lisosom.
Kantung-kantung yang terikat membran ini kaya akan enzim konsentrasi tinggi yang menghidrolisis penguraian protein dan senyawa biokimia lainnya dan ini dilepaskan saat pecahnya membran luar. Enzim tersebut adalah asam fosfatase, ribonuklease, B-glukoronidase dan protease. Karena adanya enzim pencernaan ini, lisosom disebut “kantong bunuh diri”.
Properti lain yang menarik dari lisosom, yang diisolasi dari sel ginjal adalah kemampuannya untuk menjebak dan memusatkan protein asing yang disuntikkan, menunjukkan bahwa ini dapat membantu pencernaan atau detoksifikasi bahan asing melalui pinositosis.
Kejadian:
Lisosom terdapat pada semua sel hewan kecuali sel darah merah. Mereka telah dicatat dalam jamur, Euglena, sel ujung akar jagung, kapas dan biji kacang. Pada hewan, lisosom berlimpah dalam leukosit, makrofag, sel Kupffer dan sel serupa dengan aktivitas fagositik.
Pembentukan Lisosom:
Lisosom diyakini dibentuk oleh aktivitas bersama retikulum endoplasma dan kompleks Golgi. Prekursor enzim hidrolitik sebagian besar disintesis di retikulum endoplasma kasar yang mentransfernya ke permukaan pembentuk kompleks Golgi melalui vesikelnya. Di kompleks Golgi, prekursor diubah menjadi enzim. Mereka kemudian dikemas dalam vesikel yang lebih besar yang terjepit sebagai lisosom dari wajah yang matang.
Lisosom melewati berbagai tahap dalam sel yang sama. Fenomena itu disebut polimorfisme atau keberadaan lebih dari satu bentuk morfologi. Empat bentuk lisosom yang paling berbeda diamati yaitu,
- Protolisosom primer atau protolisosom diduga berasal dari permukaan trans kompleks Golgi.
- Lisosom sekunder, heterolisosom, dan autolisosom: Yang pertama terbentuk setelah lisosom primer bergabung dengan vakuola sitoplasma yang mengandung zat ekstraseluler yang diperoleh dengan segala bentuk endositosis. Setelah fusi selesai, enzim dari lisosom primer dilepaskan ke dalam vakuola yang disebut ‘fagosom atau endosom’.
Setelah pembentukan heterolisosom atau autolisosom selesai, pencernaan enzimatik dari isi vakuolar dimulai dan seiring proses berlanjut, menjadi sulit untuk mengidentifikasi sifat lisosom sekunder dan disebut vakuola pencernaan.

- Badan residu juga disebut ‘teloly : osomes’ atau ‘badan padat’. Ini mewakili tahap akhir pemecahan bahan asing atau organel seluler oleh lisosom. Pada titik ini terjadi akumulasi bahan yang kurang tercerna yang terkait dengan membran.
Autolisis atau Autophagy:
Ini adalah penghancuran diri sel, jaringan atau organ dengan bantuan lisosom. Lisosom per pembentukan autolisis tidak melampirkan struktur yang akan dipecah. Sebaliknya mereka sendiri meledak untuk melepaskan enzim pencernaan. Autolisis terjadi pada jaringan yang mengalami degenerasi atau nekrosis.
- Sphaerosom:
Sphaerosomes (= spherosomes) adalah organel sel kecil yang dibatasi oleh membran tunggal yang berperan dalam penyimpanan dan sintesis lemak. Mereka ditemukan oleh Perner pada tahun 1953. Sphaerosomes adalah vesikel bulat dan refraktil kecil yang berdiameter 0,5-1,0 nm. Mereka muncul dari retikulum endoplasma dan dikelilingi oleh membran tunggal. 98% dari sphaerosome adalah lemak.
Protein merupakan 2% sisanya. Beberapa protein mungkin enzimatik dan mengambil bagian dalam sintesis lemak. Karena adanya lemak, sphaerosom dapat dilihat di bawah mikroskop cahaya setelah pewarnaan sel dengan pewarna Sudan dan osmium tetraoksida.
Sphaerosomes terjadi berlimpah di sel endosperm biji minyak. Sphaerosomes dari beberapa jaringan (misalnya, endosperm tembakau, ujung akar jagung) mengandung enzim hidrolitik. Oleh karena itu, mereka dianggap memiliki aktivitas lisosom.
- Badan Mikro atau Mikrosom:
Ini menunjukkan berbagai komponen seluler kecil yang dibatasi oleh membran tunggal dan nama umumnya terbatas pada organel yang memiliki flavin oksidase dan katalase. Perbedaan penting antara badan mikro hewan dan tumbuhan adalah bahwa badan mikro hewan mengandung urease, sedangkan badan mikro tumbuhan tidak memiliki enzim ini. Mikrobodi terdiri dari dua jenis — peroksisom dan glioksisom.
(i) Peroksisom (Rhodin, 1954):
Peroksisom ditemukan pada sel tumbuhan dan hewan, umumnya berhubungan erat dengan retikulum endoplasma, mitokondria, dan kloroplas. Umumnya mereka memiliki diameter 0,5-1,0 μm. Ini mengandung berbagai enzim untuk mengoksidasi berbagai substrat untuk menghasilkan hidrogen peroksida dan mengandung katalase konsentrasi tinggi untuk menghilangkan racun H 2 O 2 . Peroksisom dapat mengoksidasi substrat seperti asam urat, asam amino atau glioksilat untuk menghasilkan H,O 2 , yang segera ditindaklanjuti oleh katalase untuk direduksi menjadi oksigen dan air. Pada daun hijau, peroksisom melakukan fotorespirasi.
(ii) Glioksisom (Briedenbach, 1967):
Mikrobodi sel penyimpan lemak dalam biji minyak berkecambah ini mengandung enzim dari siklus Glyoxylate misalnya malat sintetase dan isocitrate lyase; mereka juga mengandung beberapa enzim penting dari siklus Krebs. Glioksisom umumnya terlibat dalam metabolisme trigliserida.
Seperti mikrobodi lainnya, glioksisom memiliki membran penutup tunggal dan matriks kaya enzim dengan inti kristaloid. Komponen fosfolipid dan enzim organel ini berasal dari ER. β-oksidasi asam lemak menghasilkan asetil KoA yang diubah oleh badan-badan ini menjadi asam suksinat, yang pada gilirannya, melalui jalur glikolitik terbalik menghasilkan glukosa. Kapasitas interkonversi karbohidrat lemak ini khas tanaman saja dan tidak dilaporkan pada hewan.
- Ribosom (Partikel Palade):
Ribosom ditemukan oleh Robinson dan Brown (1953) pada sel tumbuhan dan Palade (1955) pada sel hewan. Palade juga menciptakan istilah Ribosom. Mereka adalah partikel protoplasma ribonukleoprotein telanjang (RNP) dengan panjang 200-340A 0 dan diameter 170-240 A 0 yang berfungsi sebagai tempat sintesis protein atau polipeptida.
Ribosom dikenal sebagai pabrik protein. Ribosom sering membentuk susunan spiral atau roset dengan retikulum endoplasma dan tidak hanya ditemukan berasosiasi dengan RE, tetapi juga ditemukan tersebar di sitoplasma, kedua jenis ini bervariasi dalam perannya di dalam sel.
Yang pertama berlimpah ketika sintesis protein aktif untuk penggunaan sel itu sendiri (misalnya hemoglobin, enzim, dll.) dan yang terakhir ketika protein diekspor keluar sel (misalnya enzim pencernaan, antibodi, dll.)
Struktur Ribosom:
Setiap ribosom terdiri dari dua subunit yang tidak sama, berbentuk kubah yang lebih besar dan ovoid yang lebih kecil. Subunit besar memiliki tonjolan, punggungan dan tangkai. Subunit yang lebih kecil memiliki platform, celah, kepala, dan alas. Ini adalah sekitar setengah ukuran subunit yang lebih besar. Subunit yang lebih kecil cocok dengan yang lebih besar di satu ujung seperti topi. Mg 2+ diperlukan untuk mengikat dua subunit.

Ribosom dapat muncul dalam bentuk roset atau kelompok heliks yang disebut poliribosom atau polisom (Gk. poli—banyak, tubuh soma). Ribosom yang berbeda dari poliribosom terhubung dengan untaian mRNA setebal 10-20A 0 dan terbentuk selama periode sintesis protein aktif.
Tergantung pada tempat terjadinya , ribosom terdiri dari dua jenis, sitoplasma dan organel. Ribosom organel yang ditemukan dapat tetap bebas dalam matriks sitoplasma atau melekat pada retikulum endoplasma. Hanya ribosom sitoplasma bebas yang terjadi pada sel prokariotik. Ribosom terikat dan organel tidak ada.

Ukuran ribosom ditentukan oleh koefisien sedimentasi Gambar 2.28 Poliribosom pada sentrifus. Ini diukur sebagai satuan Svedberg yang disebut S (S = 1×10 13 detik). Ribosom sitoplasma prokariota adalah 70 S. Dua subunit ribosom 70 S adalah 50 S (berat molekul – 1,8 x 10 6 dalton) dan 30 S (berat molekul – 0,6 x 10 6 dalton). Ribosom sitoplasma eukariota adalah 80 S dengan dua subunit 60 S dan 40 S. Sebuah terowongan terjadi antara dua subunit untuk lewatnya mRNA.
Subunit yang lebih besar memiliki alur untuk mendorong keluar polipeptida yang baru disintesis. Ada dua situs reaktif di subunit yang lebih besar yang disebut situs-P (situs peptidil atau donor) dan situs-A (aminoasil atau situs akseptor). Kemungkinan besar kompleks aminoasil-tRNA melekat pada situs akseptor dan rantai peptida pembawa tRNA melekat pada situs peptidil atau donor.
Ribosom 80 S disintesis di dalam nukleolus. 70 S ribosom prokariota terbentuk dalam cytoplasma sedangkan organel sel semi-otonom terbentuk dalam matriksnya.
Komposisi kimia:
Secara kimiawi ribosom terdiri dari dua bagian, protein dan RNA (rRNA). Biasanya lebih banyak rRNA hadir dalam ribosom 70 S dibandingkan dengan protein (60-65: 35-40) sedangkan kebalikannya berlaku untuk ribosom 80S (40-44: 56-60). Molekul protein dan RNA dianggap disatukan baik oleh gaya elektrostatik yang membentuk ikatan garam antara gugus fosfat RNA dan gugus amino protein atau dengan membentuk kompleks magnesium atau kombinasi keduanya.

- Struktur Sitoskeletal:
Mereka adalah struktur yang sangat kecil, berserat dan berbentuk tabung yang membentuk kerangka kerja struktural di dalam sel. Mereka hanya terjadi pada sel eukariotik. Struktur sitoskeletal mempertahankan bentuk sel dan perluasannya mengatur orientasi dan distribusi organel sel. Mereka terdiri dari tiga jenis — mikrofilamen, filamen menengah, dan mikrotubulus.
(a) Mikrofilamen:
Mereka adalah batang silinder sempit panjang ultramikroskopik atau filamen protein yang terjadi pada sel tumbuhan dan hewan eukariotik. Mikrofilamen terdiri dari aktin yang merupakan 10-15% dari total protein sel.
Mereka memiliki ketebalan 6 nm dan menunjukkan penampilan manik-manik periodik karena susunan heliks yang dekat dari molekul aktin yang berbentuk bulat. Mereka juga dapat terjadi pada bundel paralel atau jaringan longgar dan umumnya terletak pada interfase sol-gel serta di bawah membran plasma. Mikrofilamen juga terhubung dengan serat gelendong, retikulum endoplasma, kloroplas dll.
Mikrofilamen bersifat kontraktil. Asosiasi dengan protein myosin tampaknya penting untuk kontraksi filamen mikro. Myofibrils serat otot juga mewakili mikrofilamen. Mikrofilamen membentuk mesin kontraktil sel, seperti pembentukan dan retraksi pseudopodia, undulasi membran plasma, mikrovili, endositosis, aliran sitoplasma, dan pergerakan organel sel lainnya.
(b) Filamen antara:
Mereka adalah filamen berongga dengan ketebalan 10 nm yang sering membentuk jaringan. Mereka terdiri dari empat jenis—-filamen Keratin—yang membentuk tonofibril dan keratin kulit; Neurofilamen yang membentuk kisi dengan kumpulan mikrotubulus di akson dan dendron sel saraf; Filamen glial adalah filamen perantara yang ditemukan dalam astrosit, dan filamen heterogen yang merupakan filamen perantara yang ditemukan di otot dan terhubung ke selubung nuklir dan sentriol, dll.
(c) Mikrotubulus:
Organel yang sangat serbaguna ini tidak hanya terlibat dalam pemeliharaan sel, tetapi juga dalam memberikan panduan arah untuk pergerakan kromosom selama pembelahan sel. Ditemukan di semua sel eukariotik, ini terutama terkait dengan silia, flagela, dan organ mirip cambuk lainnya yang terlibat dalam pergerakan.
Silinder berprotein berongga tak bercabang ini berdiameter 200-700 A 0 dan terdiri dari 13 mikrofilamen yang melintasi seluruh panjang mikrotubulus yang membungkus lumen. Penyusun utama mikrotubulus adalah glikoprotein yang disebut tubulin yang terdapat dalam dua bentuk, α-Tubulin dan β-Tubulin. Ada protein lain yang umum ditemukan yang disebut MAPS (Microtubule Associated Protein) yang fungsinya seharusnya membentuk jembatan penstabil antara mikrotubulus yang tersusun secara lateral seperti pada spindel.

- Flagela dan Silia:
Mereka adalah proses protoplasma sel yang dapat digerakkan rambut halus yang mampu menghasilkan arus dalam media cairan untuk bergerak dan lewatnya zat. Flagela lebih panjang tetapi lebih sedikit. Hanya 1-4 flagela yang terjadi per sel, misalnya banyak protista, alga motil, spermatozoa hewan, bryophyta dan pteriodophyta, dll.
Cilia lebih kecil tetapi banyak. Mereka muncul dalam kelompok ciliata protista, sel api cacing, dll. Baik silia dan flagela secara struktural serupa dan memiliki bagian yang mirip — tubuh basal, akar kecil, pelat basal, dan batang.

(i) Badan Basal atau Kinetosome:
Ini juga disebut granula basal atau blepharoplast dan terjadi tertanam di bagian luar sitoplasma di bawah membran plasma. Itu seperti silinder mikro yang memiliki struktur yang mirip dengan sentriol dengan fibril triplet yang ada di pinggiran tanpa fibril pusat, meskipun hub protein ada di sini.
(ii) Rootlet:
Mereka adalah pertumbuhan fibrilar lurik yang berkembang dari bagian bawah luar tubuh basal dan dimaksudkan untuk memberikan dukungan pada tubuh basal. Rootlets terbuat dari bundel mikrofilamen.
(iii) Pelat Dasar:
Ini adalah area dengan kepadatan tinggi yang terletak di atas tubuh basal pada tingkat membran plasma. Di daerah pelat basal, satu sub-serat dari setiap fibril perifer menghilang. Fibril pusat berkembang di daerah ini.
(iv) Poros:
Ini adalah bagian flagellum atau cilium yang menonjol seperti rambut. Panjangnya 5-10 fim untuk cilium dan 150 nm untuk flagel. Poros ditutupi di luar oleh selubung yang merupakan perpanjangan dari membran plasma. Secara internal, ini berisi matriks semi-cair yang memiliki aksonem dari 9 fibril doublet periferal dan 2 fibril singlet pusat.
Susunan ini disebut untaian 9+2 atau untai 11 dibandingkan dengan susunan sentriol atau badan basal 9+0. Dua serat singlet pusat ditutupi oleh selubung pusat protein. Mereka dihubungkan oleh jembatan ganda. Setiap fibril perifer terdiri dari dua mikrotubulus atau sub-serat B dan A.
Ia menyandang dua lengan bengkok, yang terluar memiliki pengait. Lengan samping sub-serat A terdiri dari protein dynein dengan aktivitas ATP-ase. Aktivitas semacam itu juga ada di fibril pusat. Fibril doublet perifer serta fibril singlet sentral terdiri dari tubulin. Setiap sub-serat atau fibril singlet sentral mengandung tiga belas protofilamen.
Fibril doublet perifer dihubungkan oleh penaut AB dari protein nexin antara subfiber-B dari satu dan lengan sisi dalam subfiber-A dari fibril yang berdekatan. Masing-masing sub-serat A mereka mengirimkan kolom proteinaceous radial ke pusat yang disebut berbicara. Jari-jari lebih luas secara internal untuk membentuk kepala yang terhubung ke selubung protein pusat.
Fungsi:
- Mereka membantu pergerakan pada organisme flagel akhir dan bersilia.
- Pada organisme akuatik silia dan fla gella menghasilkan arus untuk mendapatkan makanan yang disebut arus makanan.
- Pada hewan darat, silia saluran pernapasan membantu menghilangkan partikel debu di udara yang dihirup.

- Sentriol:
Sentriol adalah subsilinder mikrotubular submikroskopis kecil dengan konfigurasi sembilan fibril triplet dan kemampuan untuk membentuk duplikatnya sendiri, kutub astral dan badan basal, tanpa memiliki DNA dan penutup membran. Panjangnya sekitar 0,3 – 0,5 μm dan diameter 0,15 μm.
Biasanya dua sentriol ditemukan berasosiasi bersama dan pasangan ini sering disebut diplosome yang terletak di bagian khusus umum dari sitoplasma yang disebut sentrosfer. Centrosphere tidak memiliki organel ceil lainnya. Namun, itu mengandung bahan berserat halus. Kompleks yang terbentuk dari sentriol dan sentrosfer disebut sentrosom atau alat pusat.
Struktur:
Sebuah sentriol terbentuk dari sembilan set triplet mikrotubulus atau serat yang disusun berjarak sama dalam lingkaran di sekitar sumbu pusat imajiner yang dikenal sebagai hub. Dari luar ke dalam tiga sub-serat dari triplet fibril diberi nama C, B dan A.
Fibril triplet yang berdekatan dihubungkan oleh penghubung protein C-A. Pusat sentriol memiliki massa protein berbentuk batang yang dikenal sebagai hub. Hub memiliki diameter 2,5 nm. Dari hub, kembangkan 9 helai protein menuju fibril triplet perifer.
Mereka disebut jari-jari. Setiap spoke memiliki penebalan yang disebut X sebelum menyatu dengan sub-fiber A. Penebalan lain yang dikenal sebagai Y hadir di dekatnya. Itu melekat baik pada penebalan X maupun penghubung C-A oleh penghubung. Karena adanya jari-jari radial dan fibril perifer, sentriol memberikan tampilan roda gerobak di TS
Di bagian luar sentriol terdapat plak padat, amorf, protoplasma dalam satu atau lebih seri. Mereka disebut massa atau satelit pericentriolar. Posisi mereka ditemukan berubah dengan keadaan sel yang berbeda. Massa berfungsi sebagai pusat nukleasi untuk pertumbuhan mikrotubulus selama pembentukan aster dan produksi lebih banyak sentriol.
Fungsi:
- Sentriol memainkan peran penting dalam pembentukan gelendong dan karenanya dalam pembelahan sel.
- Mereka membentuk badan basal silia dan flagela.
- Vakuola:
Vakuola adalah area non-sitoplasma yang ada di dalam sitoplasma yang dipisahkan dari yang terakhir oleh membran spesifik. Tergantung pada isi dan fungsinya, vakuola terdiri dari empat jenis-vakuola getah, vakuola kontraktil, vakuola makanan, dan vakuola udara.
(a) Vakuola Getah:
Mereka adalah vakuola berisi cairan (getah atau getah vakuolar) yang dipisahkan dari sitoplasma oleh membran permeabel selektif yang disebut tonoplas. Sejumlah vakuola getah kecil terjadi pada sel hewan dan sel tumbuhan muda. Pada sel tumbuhan dewasa, vakuola kecil bergabung membentuk satu vakuola sentral besar. Vakuola sentral yang besar memfasilitasi pertukaran cepat antara sitoplasma dan media sekitarnya.
Getah vacuolar mengandung garam mineral, gula, asam amino, protein, produk limbah dan pigmen larut air yang disebut anthocyanin. Zat terlarut yang ada dalam getah sel mempertahankan tekanan osmotik yang tepat di dalam sel untuk turgiditas dan penyerapan airnya.
(b) Vakuola Kontraktil:
Mereka terjadi di beberapa sel protistan dan alga yang kebanyakan ditemukan di air tawar. Vakuola kontraktil memiliki membran yang sangat dapat diperpanjang dan dilipat. Itu juga terhubung ke beberapa saluran makan (misalnya Paramecium). Vakuola kontraktil mengambil bagian dalam osmoregulasi dan ekskresi.
(c) Vakuola makanan:
Mereka terjadi di sel protista protozoa, beberapa hewan tingkat rendah dan fagosit hewan tingkat tinggi. Ini dibentuk oleh fusi fagosom dan lisosom. Vakuola makanan mengandung enzim pencernaan dengan bantuan nutrisi yang dicerna. Bahan yang dicerna keluar ke sitoplasma di sekitarnya.
(d) Vakuola Udara:
(Pseudovacuoles, Gas vacuoles)—Mereka telah dilaporkan hanya pada prokariota . Vakuola udara bukanlah satu kesatuan dan juga tidak dikelilingi oleh membran umum. Ini terdiri dari sejumlah vesikel submikroskopik kecil yang masing-masing dikelilingi oleh membran protein. Vakuola udara tidak hanya menyimpan gas metabolisme tetapi juga memberikan daya apung, kekuatan mekanik, dan perlindungan dari radiasi berbahaya.
- Inklusi Sel:
Inklusi sel adalah zat tidak hidup yang ada di dalam sel. Mereka juga disebut benda ergastik. Mereka mungkin hadir dalam keadaan larut atau tidak larut dan dapat bersifat organik atau anorganik. Inklusi sel termasuk dalam tiga kategori — makanan cadangan, produk ekskretoris atau sekretoris, dan bahan mineral.
(a) Makanan Cadangan:
Mereka terdiri dari empat jenis utama—pati, glikogen, tetesan lemak, dan butiran aleuron. Butir pati terjadi pada sel tumbuhan, misalnya Beras, Oat. Butiran glikogen adalah butiran kecil penyimpanan karbohidrat yang terjadi pada sel hewan, terutama di dekat retikulum endoplasma halus di dalam hati dan sel otot.
Tetesan lemak terjadi pada sel tumbuhan dan hewan. Sel-sel hewan khusus untuk menyimpan lemak disebut adiposit. Butir aleuron mewakili protein penyimpanan yang terjadi di dalam leukoplas khusus yang disebut aleukoplas.
(b) Produk Ekskretori atau Sekretori:
Mereka dari beberapa jenis seperti lendir di beberapa sel hewan, minyak atsiri, alkaloid, resin, gusi, lateks, tanin, dll.
(c) Bahan Mineral (kristal):
Itu terjadi sebagai kristal atau kerak silika, kalsium karbonat, kalsium sulfat dan kalsium oksalat. Silika umumnya diendapkan pada sisi luar sel epidermis pada beberapa tanaman, terutama rerumputan.
Kalsium karbonat diendapkan dalam bentuk massa kristal yang disebut cystolith, misalnya dalam sel hipodermal beringin. Kalsium oksalat terjadi dalam beberapa bentuk—pasir kristal, agregat berbentuk bintang dari kristal sphaeraphide, kristal prismatik dan rafida.
- Inti:
Ditemukan oleh Robert Brown (1831) nukleus adalah bagian terpenting dari sel, karena mengontrol berbagai aktivitas metabolisme, dan juga merupakan tempat pewarisan sifat.
Umumnya sel eukariotik tidak berinti, meskipun ada juga contoh sel bi-dan multinukleat . Paramecium caudatum protistan memiliki dua inti (binukleat). Kondisi multinukleat atau polinukleat ditemukan pada beberapa sel sumsum tulang, otot lurik, pembuluh lateks, beberapa jamur dan alga.
Sel hewan atau protistan multinukleat disebut sel syncytial (misalnya, epidermis Ascaris) sedangkan pada tanaman dan jamur, mereka dikenal sebagai sel coenocytic (misalnya, Rhizopus, Vaucheria).
Ultrastruktur:
Inti tipikal interfase (fase tidak membelah atau metabolisme) berdiameter 5-25 (im. Inti ini dibedakan menjadi lima bagian—selubung inti, nukleoplasma, matriks inti, kromatin, dan nukleolus.
- Amplop Nuklir (Karyotheca):
Ini mengikat nukleus di luar dan memisahkan nukleus dari sitoplasma. Ini terdiri dari dua membran lipoprotein dan trilaminar yang disusun secara konsentrik, masing-masing setebal 60-90 A°. Membran bagian dalam halus sedangkan membran luar mungkin halus atau permukaan sitoplasmanya mengandung ribosom. Kedua membran dipisahkan oleh ruang perinuklear transparan elektron, dengan lebar 100-700 A°. Membran luar sering terhubung ke retikulum endoplasma.
Amplop nuklir dilubangi oleh pori-pori nuklir yang membantu pengangkutan material antara nukleus dan sitoplasma. Dua membran amplop menjadi kontinu di wilayah pori-pori. Pori-pori nuklir memiliki struktur yang kompleks. Mereka mungkin memiliki diafragma, septum, dan sumbat bahan padat elektron atau nukleoplasmin, annuli.
Pori inti anulasi dapat memiliki 9 silinder, satu pusat dan delapan periferal. Pori-pori nukleus memungkinkan lewatnya material secara selektif antara nukleus dan sitoplasma, misalnya K + , Na + . Cl- dan sub – unit ribosom berkumpul di nukleolus. Dengan cara yang sama RNA polimerase dan DNA polimerase meskipun disintesis dalam sitoplasma selanjutnya ditransfer ke nukleus melalui pori-pori ini.
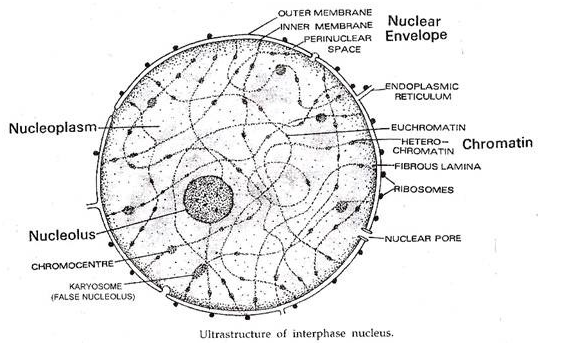
- Nukleoplasma atau Nuclear Sap atau Karyolymph:
Ini adalah zat transparan, semi-cair dan koloid yang mengisi nukleus. Ini mengandung nukleosida dan sejumlah enzim, (misalnya DNA polimerase, RNA polimerase, nukleosida fosforilase) yang diperlukan untuk sintesis dan fungsi DNA, RNA, nukleoprotein, dll.
- Matriks Nuklir:
Ini adalah jaringan serat halus protein asam yang berfungsi sebagai perancah untuk kromatin. Di pinggiran, di bawah selubung nukleus, matriks nukleus membentuk lapisan fibrosa padat yang disebut lamina nukleus di mana ujung terminal serat kromatin atau telomere tertanam.
- Kromatin:
Ini adalah kompleks fibrilar DNA-protein herediter yang dinamai demikian karena kemampuannya untuk diwarnai dengan pewarna dasar tertentu (Gk. Chroma-color; Flemming, 1879). Kromatin terjadi dalam bentuk serat halus yang tumpang tindih dan melingkar yang tampak menghasilkan jaringan yang disebut retikulum kromatin. Mereka didistribusikan ke seluruh nukleoplasma.
Serat kromatin dibedakan menjadi dua wilayah-eukromatin dan heterokromatin. Euchromatin sempit (tebal 30-80A 0 ) bernoda ringan dan bagian berserat menyebar yang membentuk sebagian besar kromatin. Heterochromatin lebih lebar (tebal 250 A 0 ) bernoda gelap dan bagian granular kental yang melekat di sana-sini pada euchromatin. Bergantung pada ukuran butiran yang dibentuk oleh heterochromatin mereka disebut chromocentres, karyosome atau false nucleoli.
Seluruh kromatin tidak berfungsi. Umumnya hanya sebagian euchromatin yang terkait dengan protein asam yang berperan dalam transkripsi atau pembentukan RNA. Selama profase pembelahan inti, serat-serat kromatin memadat membentuk sejumlah struktur seperti benang yang disebut kromosom.
- Nukleolus:
Nukleolus pertama kali ditemukan oleh Fontana pada tahun 1781, dijelaskan oleh Wagner pada tahun 1840 dan diberi nama sekarang oleh Bowman pada tahun 1840. Nukleolus adalah struktur telanjang, bulat atau sedikit tidak beraturan yang melekat pada kromatin di wilayah tertentu yang disebut wilayah pengatur nukleolus ( JUGA BUKAN).
Terkandung di dalam nukleus ukurannya tergantung pada aktivitas sintetik sel terutama pada sel yang tidak penting untuk sintesis protein, misalnya sel sperma dan blastomer. Empat jenis komponen utama dapat dilihat di dalam nukleolus, yaitu:
sebuah. Partikulat:
Terdiri terutama dari ribonukleoprotein dalam bentuk partikel kecil dengan diameter 150-200A°, dan secara struktural dan kimiawi mirip dengan ribosom sitoplasma, ini sedikit lebih kecil daripada bagian sitoplasmanya. Zona ini berada di pinggiran nukleolus.
- Bagian fibril:
Ini terdiri dari fibril protein ribonukleo dengan diameter 50-150A 0 . bahan perinokleolar. Fibril terkadang sepanjang 500A°. Bagian partikulat dan fibrillar merupakan nukleolonema.
- Bagian amorf terdiri dari protem fibril dan butiran. Bagian amorf sering disebut sebagai pars amorpha. Pada hewan, nukleolonema mungkin tampak tertanam di pars amorpha, sedangkan pars amorpha yang biasanya berada di pusat nukleolus cenderung berasosiasi dengan heterokromatin kromosom.
- Bagian kromatin adalah jaringan kusut yang menembus tubuh nukleolus tetapi melekat pada pengatur nukleolus dari kromosom . Tergantung pada posisinya, kromatin nukleolus terdiri dari dua jenis perinukleolar dan intranukleolar.
Meskipun nukleolus kaya akan protein (69-70%), RNA juga terdapat dalam jumlah yang signifikan (20-25%), menunjukkan bahwa sebagian besar RNA diproduksi di dalam nukleolus. Beberapa fraksi ribosom yang disintesis dalam sitoplasma bermigrasi ke nukleolus sebelum mengambil bentuk akhirnya. Jadi nukleolus memainkan peran penting dalam biogenesis RNA.

Kimia inti:
Dua konstituen kimia penting utama dari nukleus adalah asam nukleat, DNA dan RNA dan protein keduanya, basa dan asam. Protein dasar termasuk histon dan protamin, asam amino lisin, histidin dan arginin yang ada di keduanya.
Protein asam nukleus termasuk protein sisa kromosom. Protein asam mengandung triptofan tetapi histon tidak memiliki asam amino ini. Histon tampaknya terkait dengan kromosom sedangkan protein asam ditemukan di nukleolus dan kariolimf.
Asam nukleat merupakan 20-40% dari bahan inti, dengan DNA terutama di kromatin dan RNA didistribusikan ke seluruh inti.
Fungsi Nukleus:
- Nukleus adalah bagian integral dan esensial dari sel eukariotik dan mengandung bahan herediter yang disebut kromatin, kompleks DNA-protein. Bagian kromatin dari nukleus memiliki semua informasi genetik yang diperlukan untuk pertumbuhan dan perkembangan organisme, reproduksi, metabolisme, dan perilakunya.
- Nukleus mengontrol metabolisme sel dan aktivitas lainnya melalui pembentukan RNA (mRNA, rRNA, tRNA) yang mengontrol sintesis jenis enzim tertentu. Dengan bantuan RNA, nukleus mengarahkan sintesis protein struktural spesifik dan bahan kimia lain yang diperlukan untuk pertumbuhan sel dan pemeliharaan sel.
- Ribosom terbentuk di bagian nukleolus nukleus.
- Nukleus mengarahkan diferensiasi sel dengan memungkinkan kumpulan gen tertentu untuk beroperasi.



