Inilah catatan Anda tentang jaringan otot Tubuh Manusia!
Otot terutama dirancang untuk gerakan yang merupakan ciri khas kehidupan hewan. Salah satu sifat dasar sel hewan adalah kontraktilitas dan ini dikembangkan dalam bentuk yang sangat terspesialisasi dalam jaringan otot. Kata otot berasal dari bahasa latin musculus yang berarti tikus kecil (mus). Ini mungkin karena fakta bahwa otot-otot tertentu memiliki kemiripan dengan tikus dan tendon mewakili ekornya.

Sumber Gambar: upload.wikimedia.org/wikipedia/commons/1/10/Lateral_head_anatomy.jpg
Pada vertebrata, otot terdiri dari tiga jenis — bergaris atau sukarela, tidak bergaris atau tidak disengaja, dan jantung.
Otot bergaris terlihat lurik silang di bawah mikroskop, disuplai oleh saraf serebro-spinal dan biasanya dikendalikan secara sukarela. Oleh karena itu, mereka disebut ‘sukarela’. Namun istilah tersebut tidak sepenuhnya memuaskan. Struktur otot faring dan diafragma bergaris, tetapi tindakannya tidak sepenuhnya di bawah kendali sukarela; Otot bergaris juga disebut otot rangka atau otot somatik karena keterikatannya pada jaringan rangka. Otot-otot ini berkontraksi dengan sangat cepat, tetapi lebih mudah lelah. Otot-otot sukarela berfungsi untuk menyesuaikan organisme dengan lingkungan luarnya.
Otot yang tidak bergaris tidak menunjukkan lurik silang dan secara struktural merupakan jenis jaringan kontraktil yang paling sederhana. Mereka merespons rangsangan dengan lambat dan mampu melakukan kontraksi berkelanjutan . Otot yang tidak bergaris, juga dikenal sebagai otot polos atau visceral, tidak disengaja, karena disuplai oleh saraf otonom dan tidak berada di bawah kendali langsung kehendak. Mereka menyediakan lingkungan internal dengan kekuatan motif untuk pencernaan, sirkulasi, sekresi dan ekskresi.
Status otot jantung berada di antara otot rangka dan otot polos. Otot jantung adalah lurik silang tetapi diatur oleh saraf otonom. Mereka dikhususkan untuk memberikan kontraktilitas ritmis intrinsik jantung.
Semua otot tubuh berkembang dari mesoderm, kecuali arrectores pilorum, otot iris, dan sel mioepitel kelenjar ludah, keringat dan lakrimal yang berasal dari ektoderm.
Otot Sukarela:
Otot sukarela membentuk sekitar 42% dari total berat badan. Mereka bekerja pada sendi menghasilkan gerakan. Hanya 20% dari energi yang dibebaskan selama gerakan dinyatakan sebagai kerja, dan sisanya digunakan untuk menghasilkan panas. Ketika suhu tubuh turun di bawah normal, upaya dilakukan untuk menghasilkan lebih banyak panas dengan kontraksi otot yang cepat yang dikenal dengan istilah menggigil. Otot-otot sukarela dapat dibandingkan dengan mesin berkecepatan tinggi yang mampu mengembangkan tenaga besar, yang bekerja hanya untuk periode sedang dengan interval untuk rehabilitasi.
BAGIAN dari otot volunter-menyajikan dua bagian, berdaging dan berserat (Gbr. 7-1)
Bagian otot yang berdaging bersifat kontraktil, sangat vaskular dengan tingkat metabolisme yang lebih tinggi, dan tidak dapat menahan tekanan atau gesekan. Bagian berserat mungkin tendinous atau aponeurotik. Tendon tidak elastis, kurang vaskular dan tahan terhadap gesekan. Ketika otot menekan struktur keras, bagian berdaging digantikan oleh tendon. Jika tendon dikenakan gesekan bursa atau selubung sinovial diselingi.

Fungsi tendon:
(a) Ini memusatkan tarikan otot di tempat penyisipan.
(b) Ini sangat kuat, sehingga sepuluh don yang luas penampangnya 1 inci persegi, dapat menopang beban seberat 9.700 hingga 18.000 lbs,
(c) Serat-serat tendon dipelintir atau dianyam sehingga tarikan otot terdistribusi ke semua titik di tempat insersi.
(d) Ketika tendon terkena tarikan tiba-tiba dan tidak disengaja pada ujung insersi, tulang dapat retak tanpa ruptur tendon. Ini menunjukkan kekuatan laten yang luar biasa dalam tendon.
(e) Pada persimpangan myotendinous, serabut otot bersebelahan tetapi tidak bersambung dengan serabut tendon. Kerangka jaringan ikat otot (endo- dan perimysium) bersambung dengan kerangka serupa pada tendon. Susunan otot pada sambungan myotendinous menyerupai tampilan ‘ekor merpati’ (Gbr. 7-2).

Asal dan Penyisipan:
Setiap ujung otot dilekatkan oleh jaringan ikat ke tulang atau tulang rawan, atau struktur lainnya. Ketika otot berkontraksi, biasanya salah satu ujungnya tetap dan ujung lainnya bergerak. Dengan konvensi, ujung yang tetap disebut asal, dan yang dapat digerakkan disebut penyisipan.
Pada ekstremitas, perlekatan otot distal biasanya disebut insersi, karena bagian distal lebih dapat digerakkan. Namun, terlalu banyak tekanan tidak boleh diberikan pada origin dan insersi , karena insersi anatomi pada beberapa otot tetap terfiksasi dan origin bergerak.
Jenis insersi otot:
(1) Beberapa otot dimasukkan di dekat ujung proksimal tulang, di dekat sendi [Gbr. 7-3 (a)]. Ini meningkatkan jangkauan gerakan, tetapi kekuatan aksinya lebih kecil. Contoh-Biceps brachii, Psoas mayor
(2) Beberapa dimasukkan ke ujung distal tulang, jauh dari sendi [Gbr. 7- 3(b)]. Di sini kekuatan aksi lebih banyak tetapi jangkauan gerakan lebih sedikit. Contoh- Brachioradialis.
(3) Kadang-kadang otot dimasukkan di tengah batang tulang. Contoh- Coraco-brachialis, pronator teres.
Klasifikasi Otot Sukarela:
(A) Menurut warnanya-Otot ada dua jenis, merah dan putih. Warna tergantung pada kepadatan kapiler dan pada jumlah mio-hemoglobin dalam sarkoplasma sel otot. Pada otot merah, myo-hemo- globin lebih banyak. Otot merah dan putih menunjukkan beberapa perbedaan (Lihat Tabel):
(B) Menurut arah serat otot: otot mungkin berbentuk paralel, pennate, spiral dan cruciate.

Otot Paralel (Gbr. 7-4):
Serabut otot sejajar dengan garis tarikan. Seratnya panjang, tetapi jumlahnya relatif sedikit.
Fungsi:
(a) Kisaran gerakan lebih banyak pada jenis otot ini karena bertambahnya panjang serat.
(b) Total gaya kontraksi lebih sedikit, karena jumlah serabut yang lebih sedikit. Otot paralel dapat dibagi menjadi sub-tipe berikut:
(i) Tali otot, contoh: Sartorius, Rectus abdominis.
(ii) Otot kuadrat, contoh: Quadratus lumborum.
(iii) Otot fusiform, contoh: Biceps brachii.

Otot Pennate:
Serat berdaging miring ke garis tarikan. Seratnya pendek dan jumlah serat yang lebih banyak dapat ditampung. Otot pennate menghadirkan subtipe berikut:
(1) Unipennat [Gbr. 7-5 (a)]-Semua serat berdaging miring ke satu sisi tendon, yang terbentuk di sepanjang satu tepi otot. Ini memberikan tampilan setengah bulu.
Contoh: fleksor polisis longus, ekstensor digitorum longus, peroneus tertius.
(2) Bipennat [Gbr. 7-5(b)]-Tendon terbentuk di poros tengah otot, dan serat otot miring ke kedua sisi tendon tengah, seperti bulu utuh.
Contoh: Rectus femoris, dorsal interossei tangan dan kaki.
(3) Multipennat [Gbr. 7-5 (c)] – Serangkaian bipennat terletak berdampingan dalam satu bidang.
Contoh: Serat akromial deltoid.
(4) Cricumpennate (Gbr. 7-5 (d)]-Otot berbentuk silindris, di mana tendon sentral muncul Serat otot miring menyatu ke dalam tendon sentral dari semua sisi.
Contoh: Tibialis anterior.

Fungsi otot pennate:
(a) Jangkauan gerak berkurang karena serat otot pendek dan arah tarikan miring. Kekuatan aksi otot diselesaikan menjadi dua kekuatan komponen; satu bertindak dalam garis tarikan dan yang lainnya pada sudut kanan terhadapnya.
(b) Total kekuatan kontraksi meningkat karena jumlah serat otot yang lebih banyak.
Otot spiral:
Beberapa otot dipelintir dalam pengaturan yang dekat dengan penyisipannya. Misalnya, pektoralis mayor dimasukkan ke bibir lateral alur bicipital dengan cara berbentuk U bilaminar . Kepala klavikula pectoralis mayor membentuk lamina anterior dan kepala sternokostal diputar dari batas bawah U untuk membentuk lamina posterior.
Pengaturan spiral seperti itu membawa perlekatan otot proksimal dan distal ke bidang yang sama. Pada otot supinator, jalur spiral memberikan gerakan rotasi ke jari-jari.
Otot Cruciate:
Otot masseter dan sternocleido-mastoid termasuk dalam kategori ini, karena serat ototnya tersusun dalam bidang superfisial dan dalam yang bersilangan seperti ‘X’. Serat superfisial masseter diarahkan ke bawah dan ke belakang dari arkus zygomatic ke ramus mandibula, sedangkan serat dalam diarahkan ke bawah dan ke depan. Serabut superfisial mengangkat dan menjulurkan mandibula, dan serabut yang dalam mengangkat dan menarik mandibula. Ketika kedua set serat berkontraksi secara bersamaan, hanya elevasi yang terjadi.
(C) Menurut kekuatan tindakan- Dua jenis otot rangka ditemui, semburan dan shunt.
Pada sendi sederhana, satu tulang lebih mobile daripada tulang lainnya. Sebuah otot saat bekerja pada tulang bergerak, memberikan gaya yang menurut analisis vektor dapat diselesaikan menjadi dua komponen gaya yang tegak lurus satu sama lain – komponen ayun yang cenderung menghasilkan gerakan sudut sendi, dan komponen shunt ( trans-artikular) yang cenderung menarik tulang di sepanjang poros menuju sendi dan menekan permukaan artikular [Gbr. 7-6 (a)].
Ketika komponen ayunan lebih kuat, otot itu disebut otot sembur. Di sisi lain, dengan adanya komponen shunt yang kuat otot ditunjuk sebagai otot shunt. Pada otot yang menyembur, perlekatan tetap lebih jauh dari sendi dan perlekatan bergerak terletak dekat dengan sendi [Gbr. 7-6 (b)]. Akhirnya komponen ayunan menghasilkan semburan gerakan sudut dan komponen shunt, meskipun lemah, menjaga permukaan artikular tulang tetap berhubungan dengan sendi.
Ketika gerakan sudut melebihi 90 °, komponen shunt yang bekerja di sepanjang batang tulang mo bile cenderung mengalihkan tulang menjauh dari sendi. Brakialis adalah contoh otot semburan yang bekerja pada sendi siku.
Pada otot shunt , keterikatan tetap terletak di dekat sendi [Gbr. 7-6(c)]. Sepanjang pergerakan otot shunt, gaya tekan trans-artikular menjaga permukaan artikular tulang bergerak tetap berhubungan dengan sendi. Brachio-radialis adalah contoh otot shunt yang bekerja pada sendi siku.
Otot yang menyembur memberikan percepatan gerak sendi, sedangkan otot shunt memberikan gaya sentripetal yang menstabilkan pada sendi. Mac Conaill (1978) mengusulkan rasio partisi yang dilambangkan dengan P. Jika jarak antara sumbu sendi dan asal fungsional otot yang menyebabkan ayunan diketahui, katakanlah c, dan jarak antara sumbu sendi yang sama dan penyisipan fungsional otot seharusnya memiliki nilai, q, maka p= c / q , [Gbr. 7-6 (d)]. Bila P>1, maka otot tersebut tergolong tipe ‘spurt’, sedangkan pada kondisi sebaliknya disebut otot ‘shunt’.

Beberapa Pengamatan:
(1) Kekuatan total otot adalah jumlah kekuatan yang diberikan oleh serat-serat individualnya . Ini berbanding lurus dengan jumlah serat otot.
(2) Rentang gerak berbanding lurus dengan panjang serat otot.
(3) Kekuatan dan kecepatan gerak berhubungan dengan jarak antara titik gerak dan sumbu gerak suatu sendi. Kekuatan lebih banyak ketika jaraknya lebih jauh. Di sisi lain, kecepatan lebih besar bila jaraknya lebih pendek.
Kontraksi Otot:
Ketika otot berkontraksi untuk menghasilkan gerakan , semua serat otot tidak perlu berkontraksi secara bersamaan. Pada upaya yang lebih giat, semakin banyak serat yang terlibat. Tetapi kontraksi serat individu mana pun selalu maksimal, dan mematuhi hukum semua atau tidak sama sekali.
Saat kontraksi, bagian otot yang berdaging memendek sekitar 50 hingga 55 persen dari panjang istirahat. Jika rentang gerak otot diketahui, dapat dihitung panjang bagian berdaging dari otot paralel. Kelebihan panjang otot diubah menjadi tendon.
Otot tidak dapat berkontraksi hingga kurang dari panjang minimum tertentu. Ini dikenal sebagai insufisiensi aktif. Dalam insufisiensi pasif, otot tidak dapat diregangkan melebihi panjang tertentu tanpa cedera
Aksi Otot:
Serangkaian gerakan menghasilkan suatu tindakan. Untuk menghasilkan gerakan, kelompok otot berikut terlibat:
(a) Penggerak utama,
(b) Antagonis,
(c) Otot fiksasi ,
(d) Sinergis.
Penggerak utama:
Ini adalah otot atau sekelompok otot yang secara langsung menghasilkan gerakan yang diinginkan. Terkadang gravitasi bertindak sebagai penggerak utama. Ketika penggerak utama dari satu gerakan membantu gerakan yang berlawanan dengan perpanjangan aktif melawan gravitasi, itu dikenal sebagai tindakan paradoks. Deltoid adalah penculik sendi bahu. Ini membantu dalam adduksi selama menurunkan berat badan dari posisi horizontal. Deltoid yang berkontraksi mengontrol adduksi dengan memanjang melawan gravitasi.
Antagonis:
Otot-otot ini menentang gerakan yang diinginkan. Mereka membantu penggerak utama dengan relaksasi aktif untuk melakukan tindakan halus. Ini karena ‘Hukum persarafan timbal balik’ dan diatur oleh sumsum tulang belakang melalui refleks peregangan.
Terkadang penggerak utama dan antagonis berkontraksi secara bersamaan. Ini diatur oleh korteks serebral.
Otot fiksasi:
Ini adalah kelompok otot yang menstabilkan sendi proksimal tungkai untuk memungkinkan gerakan pada sendi distal oleh penggerak utama.
Sinergis:
Mereka adalah otot fiksasi khusus. Ketika otot melintasi dua atau lebih sendi, sinergis mencegah gerakan yang tidak diinginkan pada sendi perantara.
Selama fleksi jari dengan kontraksi otot fleksor panjang lengan bawah, sendi pergelangan tangan tetap terfiksasi oleh kontraksi ekstensor. Oleh karena itu, ekstensor pergelangan tangan bertindak sebagai sinergis selama fleksi jari.
Tulang dan Otot sebagai Sistem Pengungkit Tubuh:
Tulang dan persendian, tempat otot bekerja, berfungsi sebagai pengungkit untuk mencapai gerakan tubuh.
Untuk memahami beberapa jenis pengungkit, istilah-istilah berikut ditemui: –
(1) Titik tumpu (F) adalah titik atau garis di sekitar mana tuas bergerak, dan di dalam tubuh itu disediakan oleh sambungan.
(2) Upaya (E) mewakili gaya yang diperlukan untuk menggerakkan tuas dan merupakan titik di mana otot menyisip pada tulang untuk mengerahkan gaya kontraktilnya.
(3) Resistensi (R) adalah beban yang harus diatasi oleh kontraksi otot dan biasanya dianggap terkonsentrasi di area kecil pada tuas.
Kelas Tuas (Gbr. 7-7 a, b, c,):
Tiga kelas tuas dikenali sebagai berikut:
Pengungkit kelas satu memiliki tumpuan yang terletak di antara usaha dan perlawanan. Dalam tubuh beberapa pengungkit kelas satu ditemukan, karena pengungkit seperti itu memerlukan proyeksi pada tulang di kedua sisi sambungan. Proses olecranon ulna menerima perlekatan otot triceps, dan ketika lengan bawah diperpanjang, sendi humero-ulnar terletak di antara upaya kontraksi triceps dan resistensi yang dibentuk oleh lengan bawah dan tangan. Dengan demikian trisep bertindak untuk memperpanjang lengan menikmati penyediaan tuas kelas satu.
Pengungkit kelas dua memiliki titik tumpu di salah satu ujungnya dan hambatan berada di antara titik tumpu dan usaha. Mengangkat jari kaki adalah contoh tuas kelas dua. Upaya diterapkan di tumit, bola kaki membentuk titik tumpu dan berat badan yang terkonsentrasi di puncak lengkungan melintang merupakan perlawanan.

Pengungkit kelas tiga memiliki titik tumpu di atau dekat dengan salah satu ujung dan upaya mengintervensi antara titik tumpu dan hambatan. Ini adalah jenis tuas yang paling umum di tubuh. Biceps brachii, yang tendonnya dimasukkan ke tuberositas radial, dalam menekuk lengan bawah pada sendi siku adalah contoh nyata dari tuas kelas tiga.
Struktur Otot Sukarela:
Otot sukarela terdiri dari banyak serat silinder yang disatukan dalam matriks jaringan ikat. Serabut otot memiliki lebar yang bervariasi dari 10 µm hingga 100 µm dan panjangnya dari 1 mm hingga 5 cm. Panjang maksimum serat hingga 35 cm diisolasi dari otot sartorius. Sebagai aturan, serat otot tidak bercabang. Percabangan bagaimanapun, terjadi di otot-otot lidah.
Sitologi (Gbr. 7-8):
Setiap otot adalah sel otot individu dan terdiri dari bagian- bagian berikut:
- Sarkolema
- Sarkoplasma
- Nukleus
- Miofibril
- Miofilamen
- Mitokondria
- Retikulum sarkoplasma
- Paraplasma

Sarkolema:
Ini adalah membran sel dari serat otot, transparan, homogen dan tebal sekitar 75A. Membran terdiri dari lapisan protein luar dan dalam serta lapisan lemak di antaranya. Sarcolemma memiliki sifat listrik yang luar biasa.
Ini mempertahankan konsentrasi ion natrium dan klorida yang lebih tinggi di luar serat, dan konsentrasi ion kalium yang lebih tinggi di dalam serat. Hasil bersih dari keseimbangan ionik ini adalah perbedaan potensial sekitar -70 milivolt antara sisi dalam dan sisi luar serat otot istirahat. Ketika impuls saraf mencapai ujung saraf motorik dari serat otot, perbedaan potensial dihapuskan. Depolarisasi ini berlangsung cepat di sepanjang sarcolemma, dan serat otot berkontraksi.
Sarkoplasma:
Ini adalah sitoplasma semi-cair, non-kontraktil di mana komponen lain tertanam.
Inti:
Nuklei multipel, berbentuk oval, dan periferal terdistribusi di bawah sarkolema. Mereka terletak di sepanjang sumbu serat otot. Sebanyak beberapa ratus inti dapat hadir dalam satu serat tunggal. Oleh karena itu, setiap sel otot adalah sel berinti banyak dengan inti perifer. Di dalam embrio, inti muncul di tengah serat. Kemudian, nuklei didorong ke pinggiran, jika tidak mereka akan mengganggu kelangsungan mekanisme kontraktil serat otot.
Nukleus yang terletak di tengah terdapat pada serat intrajusal dari gelendong otot mamalia, dan pada otot vertebrata bawah.
Miofibril (Gbr. 7-8, 7-9):
Ini adalah benang paralel kontraktil dan tidak bercabang yang terletak di sepanjang sumbu panjang dari seluruh panjang serat otot. Myofibrils dapat terdistribusi secara merata, atau mereka dapat diatur dalam kelompok yang membentuk area Cohneim poligonal, yang sekarang dianggap sebagai artefak persiapan.

Di bawah mikroskop terpolarisasi, setiap miofibril muncul di sepanjang pita gelap A (anisotropik) dan pita I terang (isotropik) yang bergantian sepanjang panjangnya. Panjang setiap pita kira-kira sama. Pita gelap sangat birefringent, dan oleh karena itu disebut anisotropik. Pita cahaya tidak berubah setelah cahaya terpolarisasi dan disebut pita isotropik. Pita miofibril yang berdekatan ini disejajarkan secara melintang, memberikan serat otot tampilan lurik silang.
Setiap I-band menyajikan di tengah garis melintang gelap yang dikenal sebagai Z disc atau membran Krause. Segmen miofibril antara dua cakram Z yang berurutan, dikenal sebagai sarkomer, yang merupakan alat kontraktil dengan panjang sekitar 2-5 sore pada otot yang sedang istirahat. SarcomÂere memendek selama kontraksi serat otot. Di tengah setiap Ð-band, ada area bening yang dikenal sebagai H-band (Hensen’s band).
Bagian tengah pita-H menampilkan garis gelap tipis yang dikenal sebagai garis-M, di mana filamen miosin tebal yang menempati pita Ð (lihat nanti) dihubungkan bersama secara melintang. Protein utama M-line adalah creatine kinease, yang mengkatalisis transfer gugus fosfat dari fosfokreatin ke ADP; ini menyediakan pasokan ATP yang diperlukan untuk kontraksi otot.
Miofilamen [Gbr. 7-10(A), (B)]:
Setiap miofibril terdiri dari filamen protein yang berorientasi longitudinal, yang dikenal sebagai miofilamen. Filamen protein ini adalah elemen kontraktil utama dari otot lurik. Mikroskop elektron mengungkapkan bahwa setiap sarkomer menyajikan terutama dua jenis filamen protein, tebal dan tipis, yang terletak sejajar dengan sumbu panjang miofibril dalam pola simetris.
Filamen tipis terdiri dari aktin, tropomiosin dan troponin, sedangkan filamen tebal terutama terdiri dari miosin. Miosin dan aktin bersama-sama menghadirkan 55% dari total protein otot lurik [Gbr. 7-10(a), (b)].
Filamen miosin yang tebal hanya menempati pita A, di bagian tengah sarkomer ; panjangnya l-6pm dan lebar 15 nm. Setiap filamen menebal di M-line selama kontraksi. Setiap filamen myosin menyajikan sejumlah tonjolan seperti proyeksi lateral kepala myosin, yang tersusun berpasangan dan terhubung dengan filamen aktin selama kontraksi otot (lihat nanti). Kepala berpasangan diarahkan agak jauh dari M-line.
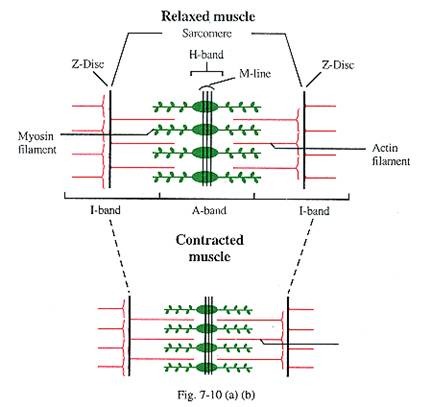
Filamen aktin tipis berada di antara dan sejajar dengan filamen miosin dan salah satu ujungnya melekat pada cakram-Z. Filamen aktin memiliki panjang 1 siang dan lebar 8 nm. Oleh karena itu, bagian dari filamen aksi terbatas pada I-band dan beberapa bagian meluas ke bagian periferal A-band. Di zona over-lapping periferal A-band, setiap filamen myosin terhubung ke enam filamen aktin dengan proyeksi lateral secara heksagonal. Namun, setiap filamen aktin dikelilingi oleh tiga filamen miosin [ Gbr. 7-10© ]

Ketika miofibril berkontraksi, filamen aktin meluncur ke dalam di antara susunan filamen miosin yang berdekatan melalui proses yang menghubungkan dan menghubungkan kembali secara berurutan antara molekul miosin dan aktin. Akibatnya, pita-I secara bertahap memendek dan menghilang, pita-H menghilang dan cakram-Z terletak di setiap sisi pita-A.
Protein filamen tebal:
Setiap filamen myosin mengandung sekitar 274 molekul myosin. Setiap molekul myosin terdiri dari enam rantai polipeptida – dua rantai berat dan empat rantai ringan. [Ara. 7-II (a), (b)].
Bagian С-terminal dari dua rantai berat myosin dipelintir bersama sepanjang bagian panjangnya untuk membentuk ekor seperti batang dari molekul myosin. Bagian yang tersisa pada Gambar 7-11 (a) ujung lain dari setiap rantai berat terlipat secara terpisah untuk membentuk proyeksi globular kepala, yang memiliki aktivitas pengikatan aktin dan aktivitas AT-Pase.

Ketika molekul myosin berkumpul untuk membentuk filamen tebal, ekor membentuk tulang punggung filamen dan kepala menonjol keluar sebagai jembatan silang. Kedua kepala molekul myosin memiliki titik perlekatan yang fleksibel antara kepala dan ekor, di mana setiap kepala dapat berayun dan berputar. Titik persimpangan kepala-ekor ini membagi molekul miosin menjadi dua sub-fragmen: meromiosin ringan (LMM) yang mewakili sebagian besar ekor, dan meromiosin berat (HMM) yang mewakili sisa ekor dan dua globular.
kepala.

Dua pasang rantai cahaya yang berbeda secara kimiawi dikaitkan dengan masing-masing kepala miosin. Mereka memediasi regulasi sensitif Ca2+ dari aktivitas ATPase myosin.
Protein filamen tipis [Gbr. 7- 12(a), (b)]:
Aktin:
Ini hadir sebagai polimer aktin berfilamen (F-aktin), yang masing-masing terdiri dari dua untaian monomer globular (G-aktin) yang saling memutar dalam formasi heliks ganda . Setiap monomer G-aktin mengandung situs pengikatan untuk myosin.
Selama polimerisasi G-aktin untuk membentuk F-aktin, mereka mengikat dari belakang ke depan (С-terminal dari satu asam amino berikatan dengan terminal-N dari asam amino berikutnya); ini memberikan polaritas yang dapat dibedakan ke filamen. Di setiap sisi cakram-Z untaian heliks dari filamen aktin penahan berputar ke arah yang berlawanan, sehingga membuat cakram-Z agak zig-zag dalam garis besarnya. Protein – α (alfa) actinin, komponen utama dari Z-disc, dianggap sebagai jangkar filamen aktin ke wilayah ini dan mengikat sarkomer yang berdekatan bersama-sama, sehingga menjaga miofibril tetap dalam register.
tropomiosin:
Setiap filamen tropomiosin adalah molekul panjang dan tipis dengan panjang sekitar 40 nm, dan terdiri dari dua rantai polipeptida yang tersusun dalam kumparan a-heliks. Filamen ini berjalan di atas 7 monomer aktin di sepanjang tepi luar alur antara dua untaian aktin yang terpuntir dan menunjukkan sedikit tumpang tindih dengan molekul tropomiosin berikutnya.
Troponin:
Ini adalah kompleks dari tiga subunit dengan panjang keseluruhan 27 nm. Subunit tersebut adalah: Troponin-1, troponin-C, dan troponin-T, Troponin-I menghambat interaksi aktin-miosin ; troponin-C mengikat ion kalsium; troponin-T melekat kuat pada satu situs spesifik pada setiap molekul tropomiosin.
Susunannya sedemikian rupa sehingga untuk setiap 7 subunit aktin terdapat satu molekul, masing-masing tropomiosin dan troponin. Molekul-molekul ini bertindak sebagai protein pengatur dalam mengendalikan kontraksi otot. (Lihat Gambar 7-12(a), (b)].


Mitokondria:
Mereka juga dikenal sebagai sarkosom dan terletak di barisan antara miofibril. Mito chondria memberikan energi untuk kerja serat otot.
Retikulum Sarkoplasma (Gbr. 7-13 (A), (B)]:
Ini adalah retikulum endoplasma permukaan halus yang mengelilingi miofibril. Retikulum terdiri dari dua jenis struktur membran yang bersentuhan satu sama lain. Jenis pertama dikenal sebagai tubulus Centro, dan yang kedua terdiri dari struktur membran yang saling berhubungan kompleks.
Setiap miofibril dikelilingi oleh sistem tubulus melingkar dan bercabang yang berasal dari pertumbuhan tubulus dari sarkolemma . Lumen tubulus terbuka di permukaan sarcolemma. Sistem tubulus Centro sebenarnya merupakan kelanjutan dari sarcolemma. Pada otot lurik amfibi, tubulus-T memanjang di setiap cakram-Z. Namun, pada otot lurik mamalia, dua tubulus T melingkari setiap sarkomer dari setiap miofibril di persimpangan pita A dan I.
Tubulus menyampaikan gelombang depolarisasi dari sarkolema ke setiap sarkomer, dan gelombang menyebar melintang melintasi serat.

Sebuah sistem struktur membran yang kompleks [Gbr. 7-13(a)]:
Sistem ini terdiri dari tiga struktur interkoneksi—-terminal cisterna, saluran longitudinal dan kantong H-band.
Cisterna terminal mengelilingi setiap sarcoma sebelum persimpangan pita A dan I, dan dikelilingi secara eksternal oleh sentrotubulus. CisÂterna mengandung bahan granular yang kaya akan ion kalsium. [Ara. 7-12(a) & 12(b)].
Saluran memanjang juga dikenal sebagai sarcotubules terletak di atas A-band, dan membentuk semacam jaringan yang menghubungkan cisterna terminal dengan kantong H-band.
Kantung H-band terletak di tengah
dari sarkomer berlawanan tingkat H-band. Dua cisterna terminal di setiap sarcomere di persimpangan pita A dan I, dan tubulus T yang berseberangan di sekitar cisterna membentuk kompleks tiga struktur membran yang dikenal sebagai triad otot.
Fungsi retikulum sarkoplasma [Gbr. 7- 14]:
(a) Selama relaksasi, kombinasi molekul troponin dan tropomiosin membentuk alat pengunci yang mencegah molekul aktin berinteraksi dengan kepala miosin pada filamen tebal yang berdekatan.

(b) Ketika gelombang depolarisasi meluas sepanjang tubulus-T, cisterna terminal retikulum sarkoplasma distimulasi untuk melepaskan ion kalsium. Ion kalsium berikatan dengan situs pengikatan Ca +2 pada troponin dan menyebabkan perubahan konformasi. Hal ini menyebabkan tropomiosin berguling lebih dalam ke alur yang dibentuk oleh dua helai heliks monomer aktin. Akibatnya, blok sterik berkurang dan ini memungkinkan interaksi aktin-miosin yang lengkap. Selama kontraksi, kepala miosin terhubung secara berurutan dan kemudian terputus dari molekul aktin di sepanjang filamen tipis, sehingga menggerakkan filamen tipis di sepanjang filamen tebal.
(c) Energi kontraksi otot berasal dari ATP yang mengikat kepala myosin dan mengembangkan afinitas tinggi untuk aktin. Ion kalsium yang terdifusi mengaktifkan ATP-ase dari kepala myosin yang dengan cepat menghidrolisis ATP untuk memutuskan kepala myosin dari molekul aktin secara berurutan, sampai filamen aktin melenyapkan kantung pita-H.
(d) Setelah itu ion kalsium masuk kembali ke kantong pita H retikulum dengan aksi pemompaan dan pemisahan ATP lebih lanjut ditahan.
Sejak generasi ATP berhenti setelah kematian, aktin dan myosin tetap terkunci bersama dalam posisi tetap, dan keadaan otot ini dikenal sebagai rigor mortis yang bertahan selama beberapa jam setelah kematian sampai autolisis terjadi.
Butiran paraplasma:
Beberapa serat otot kaya akan glikogen, lipid, dan lemak. Diperkirakan bahwa glikogen menyediakan sumber kalori yang siap pakai.
Organisasi Otot Rangka:
(1) Endomysium-lt adalah selubung halus jaringan ikat yang menutupi setiap serat otot di luar sarcolemma.
(2) Perimysium-Serat otot dikelompokkan bersama menjadi fasciculus, dan setiap fasciculus ditutupi oleh selubung jaringan ikat yang dikenal sebagai perimyÂsium.
(3) Epimysium-Seluruh otot ditutupi oleh selubung jaringan ikat yang dikenal sebagai epimysium.
Histogenesis serat otot lurik (Gbr. 7-15):
Sebagian besar otot rangka dikembangkan dari miotom mesoderm paraksial. Selama minggu kelima kehidupan embrio, sel-sel myotome menjadi berbentuk gelendong dan dikenal sebagai myoblas. Setiap myoblast menghadirkan satu nukleus dan mengalami mitosis berulang dengan kecepatan tinggi. Kemudian myoblast bergabung dari ujung ke ujung untuk membentuk myotube yang panjang, tubulus sempit yang berisi satu baris banyak inti di tengahnya.
Dalam myotubes, striasi silang muncul selama bulan kedua, oleh deposisi linier butiran protein dalam sitoplasma yang akhirnya menyatu membentuk miofibril. Pada awalnya myofibrils terletak di pinggiran myotubes, dan inti menempati posisi sentral. Dengan peningkatan jumlah myofibrils, inti didorong ke pinggiran, dan myotube diubah menjadi serat otot.
Beberapa sel satelit bernukleus ditemukan di antara membran basement dan membran plasma serat otot. Meskipun inti myotube tidak membelah, sel-sel satelit mengalami mitosis dan kemudian bergabung dengan myotube meningkatkan jumlah inti. Jadi sel-sel satelit berperilaku sebagai myoblast.

Setelah sekitar empat atau lima bulan perkembangan , massa otot individu menerima kuota penuh serat otot, dan setelah itu serat otot tidak berlipat ganda. Selanjutnya, otot meningkat dalam ukuran tetapi tidak dalam jumlah pertumbuhan serat individu.
Pertumbuhan dan Regenerasi:
Menjadi sangat terspesialisasi, otot rangka tidak beregenerasi dengan pembelahan sel dalam kondisi normal. Namun, jika bagian dari serat otot dihancurkan, regenerasi dimungkinkan. Pada vertebrata yang lebih rendah, potensi regenerasi jauh lebih besar. Hipertrofi otot setelah latihan disebabkan oleh peningkatan ukuran (bukan jumlah) serat individu.
Pasokan Vaskular Otot Sukarela:
Arteri utama dan saraf otot rangka biasanya masuk bersama di hilus neurovaskular . Di dalam substansi otot arteri bercabang di epimysium dan perimysium, bercabang menjadi arteriol dan mengeluarkan kapiler yang dibawa oleh endomisium halus.
Setiap serat otot disertai oleh satu set kapiler paralel, yang mengeluarkan cabang samping di sudut kanan serat. Otot-otot sukarela disuplai oleh pleksus kapiler yang kaya. Satu cm persegi otot disuplai oleh kapiler sepanjang sekitar 8 meter.
Metode masuknya arteri:
(a) Kadang-kadang arteri masuk pada salah satu ujung otot. Contoh. Gastrocne mius.
(b) Pada beberapa otot misalnya Biceps brachii, arteri menembus bagian tengah otot.
(c) Otot seperti adduktor magnus disuplai oleh rangkaian pembuluh anastomosis.
Pasokan limfatik:
Pembuluh limfatik otot rangka sebagian besar terbatas pada epimysium dan perimysium . Namun, limfatik tidak ada di endomisium tidak seperti otot jantung.
Pasokan Saraf Otot Sukarela:
Saraf ke otot rangka adalah saraf campuran, terdiri dari 60% serabut motorik dan 40% serabut sensorik.
Pasokan Motor (Gbr. 7-18):
(a) A-neuron (alfa) bermielin tebal mensuplai serat ekstrafusal otot yang menghasilkan gerakan.
(b) Neuron у-efferent (gamma) bermielin tipis memasok daerah kutub serat intrafusal dari gelendong otot untuk pemeliharaan tonus otot.
(c) Serabut simpatik tanpa mielin menyediakan suplai vasomotor ke pembuluh darah.
Saraf sensorik:
(1) Beberapa serabut menyampaikan sensasi nyeri dari ujung saraf bebas di sekitar serabut otot.
(2) Beberapa serabut muncul dari puscle cor lamellated di jaringan ikat.
(3) Annulo-spiral dan ujung semprotan bunga dari gelendong otot-Ini adalah reseptor peregangan dan mengatur tonus otot (Gbr. 7-18).
Titik Bermotor:
Ini adalah titik masuk batang saraf yang biasanya memasuki permukaan otot yang dalam. Stimulasi listrik otot paling efektif pada titik motorik.
Satuan Motor:
Jumlah serat otot dalam otot sadar yang disuplai oleh neuron motorik tunggal, dikenal sebagai unit motorik. Unit motor mungkin besar atau kecil.
Unit Motor Besar:
Dalam hal ini, satu neuron motorik memasok sekitar 100 hingga 200 serat otot. Otot besar dengan unit motorik besar yang lebih sedikit dapat melakukan gerakan kasar.
Unit Motor Kecil:
Ini berarti bahwa satu neuron hanya mensuplai sekitar 5 sampai 10 serat otot. Oleh karena itu, otot dengan banyak unit motorik kecil mampu melakukan tindakan yang halus dan tepat. Otot ibu jari dan bola mata memiliki unit motorik kecil.
Persimpangan Otot Neuro (Gbr. 7-16):
Akson terminal dari saraf motorik mencapai serat individu dari otot rangka sebagai ujung khusus yang mungkin terdiri dari dua tipe- motor end plate atau terminal ‘en plaque’ dan ujung jejak atau terminal ‘en grape’. Pelat ujung motorik ditemukan pada otot cepat yang mengalami kontraksi fasik dan tampak seperti perluasan seperti cakram di dekat bagian tengah serat otot. Ujung jejak muncul di otot lambat untuk kontraksi tonik dan dimanifestasikan oleh kelompok ekspansi seperti anggur di sepanjang serat otot. Pada kedua jenis, prinsipnya sama karena sambungan neuro-otot adalah sinapsis antara sel saraf dan sel otot.

Pelat ujung motor terdiri dari kumpulan sarkoplasma granular lokal yang dikenal sebagai sol yang dapat menonjol dari permukaan di bawah sarkolema (Gbr. 7-16). Satu-satunya berisi sejumlah inti besar dan mitokondria banyak dan sarcolemma menyajikan depresi kecil yang dikenal sebagai lipatan atau selokan junctional. Akson yang mendekat dari saraf motorik kehilangan selubung mielinnya dan terbagi menjadi sejumlah cabang terminal yang menempati palung atau selokan telapak kaki. Aksoplasma terminal saraf dan sarkoplasma telapak kaki tidak langsung bersambung. Hubungan antara keduanya adalah salah satu kontak permukaan antara axolemma dan sarcolemma, dipisahkan oleh interval sekitar 200A hingga 300A yang mengandung bahan amorf yang berasal dari membran dasar dan selubung sel akson dan otot.
Endoneurium saraf bersambung dengan endomisium otot. Kadang-kadang, sel-sel teloglia yang berasal dari sel-sel Schwann cenderung di sepanjang terminal saraf dan menutupi palung seperti penutup. Di sepanjang palung, sarcolemma menghadirkan serangkaian lipatan paralel dengan pola palisade. Ini dikenal sebagai aparatus subneural yang kaya akan kolinesterase. Aksoplasma terminal saraf mengandung mitokondria dan sejumlah vesikel presinaptik jernih elektron yang membawa asetilkolin.
Selama konduksi impuls saraf, asetilkolin dibebaskan di ujung pelat dan bertindak sebagai pemancar impuls secara kimiawi.
Mekanisme Transmisi Neuro-otot:
(a) Vesikel presinaptik terminal saraf mengandung asetilkolin yang dilepaskan dari aksoplasma, ketika vesikel melebur dengan membran aksolemmal. Ion kalsium mendorong pelepasan asetilkolin, dan magnesium mengantagoniskannya.
(b) Asetilkolin berdifusi dengan cepat melintasi celah kecil antara axolemma dan sarcolemma dari motor end plate, dan bergabung dengan reseptor khusus pada membran sarcolemmal postsynaptic .
Akibatnya, permeabilitas sar colemma terhadap berbagai ion meningkat, khususnya terhadap natrium dan kalium. Saat permeabilitas membran meningkat, potensial membran turun, sehingga perbedaan potensial sekitar -70 milivolt antara permukaan dalam dan luar sarkolema serat otot yang beristirahat dihapuskan. Gelombang depolarisasi berkembang dengan cepat di sepanjang sarkolema, mencapai cakram-Z sarkomer individu melalui sentrotubulus dan retikulum sarkoplasma, dan serat otot berkontraksi.
Obat tubo-curarine mencegah kombinasi asetilkolin dengan reseptor khusus di motor end plate dengan persaingan substrat, dan memblokir transmisi neuro-otot. Konsentrasi asam laktat yang tinggi selama latihan otot yang berkepanjangan dapat menyebabkan kelelahan akibat blok parsial pada transmisi neuromuskuler.

(c) Akhirnya, asetilkolin dihancurkan oleh enzim spesifik kolinesterase, yang kaya akan aparatus subneural dan memecah asetilkolin menjadi kolin dan asam asetat. Kolin membantu sintesis lebih banyak asetilkolin.
Spindel Neuro-otot (Gbr. 7-17):
Ini adalah enkapsulasi berbentuk gelendong, organ reseptor sensorik khusus yang ditempatkan secara longitudinal di antara fasikulus serat ekstra-fusal otot volunter. Spindle otot berkaitan dengan pemeliharaan tonus otot, dan mereka ditemukan di sebagian besar
otot rangka umumnya di dekat persimpangan myo-tendinous. Gelendong bertindak sebagai reseptor peregangan, dan otot-otot kecil tangan dan kaki diberkahi dengan lebih banyak gelendong.
Setiap spindel, rata-rata, berukuran sekitar 2mm hingga 4mm. panjang dan terdiri dari kapsul berserat yang berisi sekitar 2 sampai 14 serat otot intra-fusal.
Serat intra-fusal terdiri dari dua jenis, kantong bening dan. serat rantai inti. Kedua jenis serat tersebut memiliki daerah ekuator non-kontraktil tanpa striasi dan daerah kutub lurik kontraktil. Namun, jumlah kedua jenis serat tersebut bervariasi dalam spindel yang berbeda. Serat kantong nukleus lebih panjang dan lebih besar dari jenis lainnya dan terdapat perluasan di daerah khatulistiwa yang dikenal sebagai kantong nukleus yang berisi kumpulan nukleus numerous. Selain itu, ujung perifer dari serat tersebut melampaui kapsul gelendong dan melekat pada perimysium dari serat ekstra-fusal atau ke tendon yang berdekatan.
Serabut rantai nukleus lebih pendek dan sempit, terkurung di dalam kapsul spindel, dan daerah ekuatornya mengandung rantai nuklei dalam satu baris. Ujung kutub dari serat tersebut melekat pada kapsul atau pada selubung serat kantong nukleus. Serat tas nuklir tidak memiliki garis-M dan berkontraksi perlahan, sedangkan serat rantai nuklir memiliki garis-M dan menunjukkan kedutan yang lebih cepat. Serat intrafusal individu diselimuti oleh selubung aksial bagian dalam, dan interval antara kapsul spindel luar dan selubung bagian dalam yang disebutkan ditempati oleh getah bening yang kaya akan asam hialuronat.
Serabut intra-fusal dipersarafi oleh saraf sensorik dan motorik. Terminal saraf sensorik terdiri dari dua jenis, ujung annulo-spiral (primer), dan ujung bunga-spray (sekunder). Ujung annulo-spiral mengelilingi daerah ekuator dari kantong nukleus dan serat rantai nukleus, dan berasal dari serat aferen grup IA yang bermielin tebal.
Ujung semburan bunga membungkus serat rantai nuklir saja, dan didistribusikan pada satu atau kedua sisi di luar daerah khatulistiwa; ujung seperti itu milik serat aferen kelompok II. Kedua jenis ujung sensorik bertindak sebagai reseptor peregangan; reseptor distimulasi ketika serabut intrafusal diregangkan secara pasif selama relaksasi serabut ekstrafusal seluruh otot (karena serabut intrafusal sejajar dengan serabut ekstrafusal), atau ketika daerah kutub intrafusal serat berkontraksi secara aktif. Mungkin reseptor peregangan merespons lebih efektif saat diregangkan secara pasif.
Serat motorik juga dikenal sebagai, serat fusimotor memasok daerah kutub otot intra-fusal dan sebagian besar berasal dari neuron у-efferent dari kolom abu-abu anterior sumsum tulang belakang. Neuron γ-eferen terdiri dari dua jenis, γ l dan γ 2. Serabut γl terminasi sebagai ujung lempeng di dekat ujung serat intrafusal, dan neuron γ2 membentuk ujung jejak yang dekat dengan daerah berinti. Beberapa kali kolateral dari neuron β-efferent memasok, selain serat ekstra-fusal, kutub ekstrim serat intra-fusal sebagai ujung pelat. Serat fusimotor, ketika dirangsang, menghasilkan kontraksi daerah kutub, dan akhirnya reseptor peregangan daerah khatulistiwa dirangsang. Secara fungsional ada dua jenis serat fusimotor, dinamis dan statis.
Serabut fusimotor dinamis memasok otot intra-fusal jenis ‘kantong nuklir’ dan berkaitan dengan pemantauan posisi dan indra kecepatan dalam otot yang berubah dengan cepat. Serat fusimotor statis memasok serat ‘rantai nuklir’ dan merekam perubahan tersebut di atas selama adaptasi otot yang lambat. Bukti terbaru menunjukkan bahwa neuron gamma memantau tingkat kontraksi otot melalui loop refleks gamma dan menarik perbandingan antara gerakan yang dimaksudkan dan gerakan yang sebenarnya.
Nada Otot (Gbr. 7-18):
Ini berarti keadaan kontraksi sebagian otot untuk mempertahankan panjang otot yang konstan, ketika suatu gaya diterapkan untuk memanjangkan otot. Oleh karena itu, otot tidak sepenuhnya rileks bahkan dalam kondisi istirahat. Nada otot dipertahankan oleh busur refleks monosinaptik yang dikenal sebagai refleks peregangan.
Regulasi Tonus Otot:
(a) Ketika serabut ekstra-fusal dari seluruh otot berelaksasi, daerah ekuatorial dari serabut intra-fusal dari gelendong otot diregangkan, dan setelah itu ujung annulo-spiral dan flower-spray distimulasi. Impuls aferen yang dihasilkan, mencapai sumsum tulang belakang melalui sel pseudo-unipolar dari ganglia akar dorsal dan membentuk relai monosinaptik dengan neuron α- dari sel tanduk anterior. A-neuron, pada gilirannya, menembaki serat ekstra-fusal dan mempertahankan panjang otot yang konstan. Oleh karena itu, tonus otot meningkat saat istirahat dan berkurang selama aktivitas otot.
(b) Pada stimulasi neuron у-efferent dari sel tanduk anterior, daerah kutub serat intra-fusal gelendong mengalami kontraksi independen dan daerah ekuator diregangkan. Refleks peregangan monosinaptik kemudian terjadi, dan serabut ekstra-fusal dibawa ke dalam kontraksi oleh stimulasi a-neuron.
Aktivitas neuron-y diatur oleh sistem ekstra-piramidal , khususnya oleh traktus retikulospinal yang membawa serat fasilitator atau penghambat.
Fenomena Orbeli:
Otot lurik seperti diafragma disuplai oleh saraf somatik dan simpatik. Yang terakhir selain pembuluh darah, memasok otot lurik. Disarankan bahwa stimulasi saraf simpatik menunda kelelahan pada otot yang dipaksa untuk berkontraksi dengan penembakan berulang saraf motorik somatik.

Pendidikan Ulang Motorik:
Kadang-kadang transplantasi otot digunakan pada manusia untuk mengatasi kelumpuhan. Pada kelumpuhan ekstensor pergelangan tangan, tendon fleksor melekat pada dorsum tangan untuk menggantikan tangan yang lumpuh. Dengan pendidikan ulang otot yang tepat, orang tersebut mampu menghasilkan ekstensi pergelangan tangan dengan otot yang dialihkan, terkadang dengan akurasi yang mengagumkan. Hal ini dapat dijelaskan oleh fakta bahwa sumsum tulang belakang memiliki pola saraf untuk otot individu atau kelompok otot, sedangkan korteks serebral hanya berkaitan dengan gerakan dan bukan otot individu yang mempengaruhi mereka.
Bursa Sinovial dan Selubung Tendon:
Bursa adalah kantung tertutup berisi cairan pelumas dan menyerupai ukuran dan bentuk koin. Strukturnya mirip dengan membran sinovial, dan dibedakan dari jaringan fibro-areolar. Ketika tendon meluncur di atas tulang atau ligamen, bursa tampaknya mengurangi gesekan dan memungkinkan gerakan bebas. Bursae lebih umum terjadi di sekitar sendi sinovial, dan terkadang berkomunikasi dengan rongga sendi dengan menusuk kapsul fibrosa. Bursa diperlukan saat tendon atau tulang mengalami gesekan pada satu permukaan saja.
Jenis Bursae:
(a) Bursa subtendinous:
Intervensi antara tendon dan tulang, tendon dan ligamen, atau antara dua tendon yang berdekatan. Sebagian besar bursa anggota badan termasuk jenis ini.
(b) Bursa artikular:
Ini melayani fungsi sendi. Bursa yang mengintervensi antara dens of axis dan ligamen transversal atlas adalah contoh dari varietas ini.
(c) Bursa subkutan:
Tampak antara kulit dan tonjolan tulang. Contoh-Bursa infra-patellar subkutan (bursa pembantu rumah tangga).
Selubung Sinovial Tendon (Gbr. 7-19): AAAA:
Ketika tendon melewati kanal osseo-fibrous, itu diselimuti oleh bursa tubular bilaminar yang dikenal sebagai selubung sinovial. Lapisan luar atau parietal dari selubung melapisi saluran fibrosa osseo, dan lapisan dalam atau visceral menutupi tendon. Kedua lapisan tersebut bersambung satu sama lain pada ujung-ujungnya . Selain itu, kedua lapisan tersebut bersambung sepanjang sumbu panjang tendon membentuk lipatan sinovial yang dikenal sebagai mesotendon, yang menyediakan pembuluh darah ke tendon. Refleksi mesotendon terjadi di sepanjang permukaan yang paling tidak terkena tekanan. Untuk meningkatkan jangkauan pergerakan tendon, kadang-kadang mesotendon menghilang sebagian besar dan diwakili oleh benang yang dikenal sebagai vincula tendinum.

Vinculae hadir di fleksor sepuluh jari. Bukan hal yang aneh untuk menemukan lebih dari satu tendon yang diselubungi oleh satu selubung sinovial.
Urgensi yang bertanggung jawab atas munculnya selubung sinovial sedemikian rupa sehingga tendon harus mengalami gesekan pada dua permukaan atau lebih. Untuk memudahkan pelumasan, selubung sinovial memanjang sekitar 1 cm. di kedua sisi situs gesekan.
Secara perkembangan, ketika tendon tumbuh dalam jaringan ikat yang tidak berdiferensiasi, sel-sel mes enchymal disusun secara konsentris di sekitarnya. Kemudian sel-sel ini terpisah membentuk rongga selubung sinovial, yang dilapisi oleh mesothelium.
Otot Tidak Bergaris atau Tidak Disengaja:
Otot yang tidak bergaris atau polos tersebar luas di dalam tubuh, dan menjaga keseimbangan fisiologis normal dengan memberikan kekuatan motif untuk pencernaan, sirkulasi, sekresi, dan ekskresi. Mereka hadir di dinding visera tubular dan sakular berongga, saluran kelenjar eksokrin, pembuluh darah, pohon trakeo-bronkial, di stroma organ padat, iris dan tubuh ciliary, di arrectores pilorum dan kelenjar keringat kulit.
Perkembangan:
Otot tidak bergaris berkembang dari mesoderm splanchnic, kecuali otot iris dan arrectores pilorum kulit yang berasal dari ektodermal.
Struktur (Gbr. 7-20):

Serabut otot terdiri dari sel-sel berbentuk spindel yang memanjang dengan nukleus oval di tengah, dengan panjang rata-rata 40 µm hingga 80 µm. Sitoplasma mengandung miofibril yang lurik longitudinal tetapi lurik silangnya mencolok karena tidak ada. Mitokondria hadir di antara miofibril dan setiap miofibril mengandung filamen aktin yang terletak secara longitudinal. Keberadaan filamen miosin yang lebih tebal pada otot polos masih diperdebatkan. Beberapa pekerja mengamati molekul yang lebih kecil dari filamen miosin setelah menggunakan fiksatif khusus yang mungkin menunjukkan hubungan silang antara filamen aktin dan miosin.
Susunan Otot Tak Sadar:
Dalam jeroan berongga, otot-otot yang tidak bergaris tersusun dalam lembaran atau lapisan, yang disatukan oleh jaringan areolar. Disposisi serat diatur sesuai dengan fungsi jeroan dan pengaturan berikut dapat diamati:
(a) Pembuluh darah sering membutuhkan perubahan ukuran lumen untuk menyesuaikan aliran darah dengan penyempitan atau dilatasi dinding pembuluh darah. Oleh karena itu, serat otot dindingnya sebagian besar diatur secara melingkar.
(b) Untuk mendorong isi sepanjang visera tubular hanya dalam satu arah, otot-otot dibedakan terutama menjadi dua lapisan, memanjang dan melingkar. Jenis gerakan ke bawah di sepanjang tabung ini dikenal sebagai peristaltik, yang biasanya diamati di sepanjang saluran pencernaan, di ureter dan saluran rahim. Otot sirkular menyempitkan tuba untuk mencegah aliran balik, sedangkan otot longitudinal memperpendek dan melebarkan tuba distal ke konstriksi untuk menampung isi yang bergerak maju.
Mekanisme gerak peristaltik di usus (Gbr. 7-21):
Otot polos usus halus tersusun dalam lapisan bujur luar dinal dan sirkular dalam. Selama peristaltik gelombang penyempitan, yang disebabkan oleh kontraksi otot melingkar, meluas ke bawah sepanjang usus dan didahului oleh dilatasi. Carey telah mengamati bahwa otot-otot longitudinal dan sirkular dari usus kecil menjalani jalur yang agak spiral dengan cara helicoidal tangan kiri.
Otot melingkar membentuk spiral yang dekat dan berputar penuh dalam jarak 0,5 hingga 1 mm. Otot longitudinal, bagaimanapun, diatur dalam spiral terbuka membuat putaran penuh setiap 50 cm. Ketika gelombang kontraksi mulai dari titik tertentu memanjang di sepanjang kedua lapisan otot, otot longitudinal berkontraksi pada jarak yang lebih jauh dalam waktu tertentu daripada otot sirkular.
Otot longitudinal memperpendek dan melebarkan tabung, sedangkan otot melingkar menyempitkan usus. Hal ini menunjukkan bahwa setiap gelombang penyempitan harus didahului oleh gelombang dilatasi. Dilatasi usus menginduksi refleks regang dengan melebarkan spiral dekat otot sirkular, sehingga gelombang konstriksi selanjutnya dapat meluas ke distal secara efektif.

(c) Dalam jeroan sakular di mana gerakannya rumit, otot-ototnya terdistribusi secara tidak teratur menjadi tiga lapisan yang tidak jelas. Otot-otot rahim tersusun menjadi lapisan longitudinal luar, sirkular tengah, dan retikuler dalam; setiap lapisan berkaitan dengan fungsi yang terpisah. Otot longitudinal membantu mengevakuasi isi rahim terutama pada akhir kehamilan; otot sirkular mempertahankan hasil konsepsi sampai jangka penuh tercapai, sedangkan lapisan otot retikuler menyempitkan pembuluh darah dan membantu dalam hemostasis selama pemisahan afterbirths atau selama fase perdarahan dari siklus menstruasi.
Pasokan Saraf Otot Tidak Bergaris:
Otot yang tidak bergaris disuplai oleh kedua sayap sistem saraf otonom, simpatetik dan parasimpatis; satu dimaksudkan untuk kontraksi dan yang lainnya untuk relaksasi otot. Kedua sistem mengandung komponen motorik dan sensorik. Serabut saraf membentuk pleksus, sering dicegat oleh sel ganglion, dan akhirnya mencapai sel otot sebagai serat post-ganÂglionik dan non-myelinated. Perlu diperhatikan bahwa otot yang tidak bergaris tetap berkontraksi secara otomatis dan spontan, bahkan ketika terputus dari suplai sarafnya.
Dalam hubungan ini, persarafan usus patut mendapat perhatian khusus. Pasokan saraf usus terdiri dari dua bagian, ekstrinsik dan intrinsik (Gbr. 7-22).

Saraf ekstrinsik termasuk serat vagal dan simpatetik. Serat vagal setelah memasuki dinding usus, membuat sambungan sinaptik dengan sel ganglion pleksus Meissner dan AuÂerbach. Serabut simpatis adalah neuron post-ganglionik yang berasal dari ganglion seliaka dan tidak masuk ke sambungan sinaptik dengan sel pleksus ini, tetapi berakhir langsung di sel otot polos dan di dinding pembuluh darah.
Saraf intrinsik terdiri dari pleksus Auerbach dan Meissner; yang pertama terletak di antara lapisan otot melingkar dan memanjang dan yang terakhir terletak di lapisan submukosa usus. Setiap pleksus terdiri dari kelompok sel saraf dan kumpulan serat saraf. Sel-sel saraf bertindak sebagai neuron postganglionik hanya untuk serabut vagal. Jaringan saraf intrinsik ini bertanggung jawab atas gerakan spontan saluran usus, bahkan setelah suplai saraf ekstrinsik dipotong. Tetapi pergerakan usus berhenti jika pleksus intrinsik terganggu oleh aplikasi nikotin.
Pleksus Auerbach mengatur arah kontraksi gelombang peristaltik. Beberapa ahli mengklaim bahwa pleksus intrinsik mewakili busur refleks lengkap. Akson sel ganglion terbagi menjadi dua cabang. Satu cabang menerima rangsangan sensorik dari mukosa usus dan meneruskan impuls ke cabang lain tanpa melewati badan sel. Impuls kemudian mencapai otot, pembuluh darah dan kelenjar usus dan menghasilkan aktivasi motorik. Pasokan saraf ekstrinsik mengatur mekanisme neuro-otot intrinsik yang menentukan pergerakan saluran pencernaan.
Baru-baru ini diamati bahwa badan sel dari neuron pleksus Meissner bertindak sebagai neuron sensorik bipolar; serat bagian dalam mereka menyampaikan rasa distensi dari selaput lendir dan serat bagian luar membuat relai sinapik dengan dua set neuron motorik di pleksus Auerbach. Satu set (paling umum) terdiri dari serat kolinergik yang pada stimulasi meningkatkan aktivitas peristaltik dan sekretomotor kelenjar. Set lainnya melibatkan neuron peptidergik yang pada stimulasi menghambat peristaltik dan aktivitas sekretomotor dengan melepaskan zat mirip ATP.
Transmisi Gelombang Eksitasi:
Menurut transmisi eksitasi, otot yang tidak bergaris diklasifikasikan menjadi dua kelompok fungsional – tipe multi-unit dan tipe unit tunggal.
Tipe multi-unit ditemukan di lapisan otot vas-deferens. Pada tipe ini, serabut otot polos benar-benar terpisah satu sama lain, dan terdapat ujung saraf untuk setiap sel otot. Saat impuls saraf tiba, semua serabut otot di area tertentu berkontraksi secara bersamaan.
Tipe unit tunggal juga dikenal sebagai tipe visceral ditemukan di usus. Impuls saraf yang tiba di satu sel otot, dihantarkan dengan cepat dari sel ini ke sel lainnya sehingga semuanya berkontraksi kurang lebih secara bersamaan. Gelombang eksitasi ditransmisikan oleh tarikan mekanis dari sel ke sel melalui membran sel yang menyatu atau jembatan protoplasma antar sel.
Pertumbuhan dan Regenerasi:
Biasanya otot yang tidak bergaris tidak berkembang biak dengan mitosis. Hipertrofi fisiologis otot-otot rahim selama kehamilan disebabkan oleh pembesaran serat individu, meskipun beberapa pihak berwenang mengklaim bahwa beberapa sel meningkat jumlahnya (hiperplasia) dengan mitosis dari sel mesenkim yang tidak berdiferensiasi.
Sel mio-epitel:
Ini adalah sel epitel berbentuk gelendong yang memiliki sifat kontraktil. Sel-sel myoepithelial dikembangkan dari ektoderm dan ditemukan di kelenjar lakrimal, salia, keringat dan susu. Sel-sel terjepit di antara membran dasar dan epitel permukaan asinus kelenjar. Mikroskop elektron menunjukkan adanya filamen aktin di dalam sel mioepitel; banyak desmosom menghubungkan sel-sel ini dengan sel-sel jaringan yang berdekatan.
Otot Jantung
Struktur:
Otot jantung atau myocardium terbatas hanya pada jantung dan akar pembuluh besar yang melekat pada jantung.
Setiap serabut otot menunjukkan persilangan, nukleus yang terletak di tengah, disk interkalasi, dan cabang samping yang sering membentuk hubungan silang dengan serabut yang berdekatan [Gbr. 7-23 (a)]. Lurik silang kurang berbeda dibandingkan dengan otot sukarela. Fitur-fitur ini, seperti yang ditemukan di bawah mikroskop cahaya, menunjukkan bahwa otot jantung membentuk syncytium tanpa gangguan tanpa dinding sel yang jelas.

Tetapi dengan bantuan mikroskop elektron, sekarang ditetapkan bahwa setiap serat otot jantung terdiri dari sejumlah sel silinder pendek yang bergabung dari ujung ke ujung, dan memberikan cabang samping untuk terhubung dengan serat yang berdekatan. Setiap sel otot memiliki sarkolema halus dan biasanya berisi satu nukleus sentral. Sarkoplasma mengandung miofibril kecuali di sekitar nukleus, aparatus Golgi, glikogen, tetesan lemak dan beberapa butiran pigmen.
Myofibrils menunjukkan cakram Ð-band, I-band, H-band dan Z yang mirip dengan otot sadar. Akun ini untuk cross-striations otot jantung. Mitokondria banyak di antara miofibril. Mereka bertindak sebagai ‘pembangkit listrik’ sel dengan memasok ATP yang memberikan energi ke otot jantung. Susunan retikulum koplasma sara mirip dengan otot lurik volunter.
Disk interkalasi adalah pita transversal yang sangat bernoda yang muncul secara berkala sekitar 70 µm di sepanjang serat. Setiap cakram memanjang di seluruh lebar
serat, dan juga memengaruhi cabang samping. Cakram lebih menonjol pada usia lanjut. Sel-sel otot individu bergabung dari ujung ke ujung dengan cakram interkalasi yang melintasi miofibril pada cakram Z. Dalam kebanyakan kasus, cakram berjalan sebagai garis lurus melintasi serat. Kadang-kadang mereka diatur secara bertahap, ketika cakram Z dari miofibril yang berdekatan tidak terdaftar satu sama lain. Setiap cakram dibentuk oleh aposisi membran sel sel otot dan bahan padat elektron pada sisi sitoplasma membran sel [Gambar. 7- 23 (b)]. Membran sel dipisahkan oleh interval sekitar 20 nm.
Persimpangan dari cakram interkalasi penting dalam kaitannya dengan transmisi gelombang kontraksi selama siklus jantung. Mungkin impuls eksitasi menyebar dari sel ke sel yang serupa dengan jenis otot tak bergaris satu unit. Ciri paling khas dari otot jantung adalah interkoneksi yang sering dilakukan oleh cabang samping. Ini melayani dasar anatomis untuk penyebaran kontraksi ke seluruh jantung.
Oleh karena itu, dosis otot jantung tidak membentuk syncytium sejati karena setiap serat otot terdiri dari sambungan ujung ke ujung dari sel-sel otot individu. Miosit jantung terdiri dari empat jenis — nodal, transisi, Purkinje dan miosit biasa. Miosit nodal dan transisional sangat tereksitasi tetapi kecepatan konduksinya lebih rendah; Serabut Purkinje memiliki kecepatan konduksi yang lebih tinggi tetapi kurang dapat dirangsang.
Sistem penghantar Jantung (Gbr. 7 – 24):
Otot jantung secara khusus dibedakan dalam sistem konduksi jantung dengan agregasi serat nodal, transisi dan Purkinje.
Serabut nodal hadir dalam nodus sinu-atrial dan atrioventrikular ‘, yang pertama bertindak sebagai ‘pembuat kecepatan’ jantung dengan mengatur kontraksi ritmis intrinsik otot jantung. Ritme alat pacu jantung, bagaimanapun, diatur oleh impuls saraf dari sistem saraf otonom. Disarankan bahwa gelombang kontraksi berasal dari miogenik daripada neurogenik, karena pada embrio serabut otot jantung mulai berkontraksi secara ritmis sebelum serabut saraf mencapai jantung.

Serabut nodal berbentuk fusiform, lurik silang kurang jelas dan lebih kecil dari serabut otot jantung biasa. Serabut-serabut itu bercabang banyak dan beranastomosis dengan cara pleksiform. Mereka tertanam dalam jaringan ikat padat, yang diresapi dengan kapiler. Ganglia perifer dari sistem parasimpatis banyak terdapat di sekitar nodus. Setiap node dipasok oleh kedua divisi sistem saraf otonom . Stimulasi saraf parasimpatis mengurangi denyut jantung, sedangkan stimulasi simpatis meningkatkannya.
Serabut Purkinje hadir di pinggiran nodus sinu-atrial dan atrio-ventrikular dan di sepanjang cabang terminal dari dua fasikuli bundel atrio-ventrikular, yang bercabang secara sub-endokardial sebagai pleksus di dinding ventrikel, dan berlanjut dengan serat miokard biasa. Serat-serat ini menghubungkan nodus SA dan nodus AV melalui traktus internodal anterior, tengah, dan posterior. Serat Purkinje berkaitan dengan kecepatan konduksi yang lebih tinggi dan semakin besar diameternya, semakin cepat pula laju konduksinya. Secara struktural, serabut Purkinje lebih besar, lebih pucat, dan tidak terlalu lurik dibandingkan serabut otot jantung biasa. Myofibrils di setiap serat dibuang di pinggiran dan inti pusat ditempati oleh glikogen.
Impuls kontraksi dimulai pada nodus sinu-atrial, mengaktifkan otot-otot atrium dan dengan demikian disampaikan ke nodus atrio-ventrikular melalui saluran internodal. Bundel atrio-ventrikular, kedua fasikulusnya dan serabut Purkinje menghantarkan impuls dari nodus atrioventrikular ke miokardium ventrikel.
Sinu-Atrial Node [Gbr. 7-24(b)]:
Ini pertama kali dideskripsikan oleh Keith dan Flack pada tahun 1907. Nodus SA berbentuk tapal kuda pada penampang melintang, terletak di bagian atas sulcus terminalis atrium kanan dan memanjang secara medis di depan muara vena cava superior. Itu jatuh sedikit lebih pendek dari epikardium, dan endokardium, dan berukuran sekitar 15 mm. panjangnya, 5 mm. dengan lebar dan 1,5 mm. dalam ketebalan.

Sebuah arteri nodal berjalan secara longitudinal melalui pusat nodus dan serat nodal, transisional dan Purkinje disusun sejajar dengan arteri dari dalam ke luar. Di pinggiran nodus, serabut Purkinje bersambungan dengan otot atrium.
Nodus Atrio-Ventrikuler:
Ditemukan oleh Tawara pada tahun 1906. Nodus AV terletak di bawah endokardium atrium kanan di bagian bawah septum interatrium dan di atas muara sinus koroner. Itu sekitar 6 mm. panjangnya, 3 mm. dengan lebar dan 1 mm. dalam ketebalan.
Nodus AV pada dasarnya terdiri dari serat transisi di tengah dan serat Purkinje di pinggiran. Serabut Pukinje menyambung dengan otot atrium di salah satu ujungnya, dan dengan bundel atrio-ventrikular di ujung lainnya.
Bundel Atrio-Ventrikuler:
Dari nodus mula-mula bundel AV berjalan ke atas di trigonum fibrosum dextrum, dan mencapai tepi posterior bagian membran septum ventrikel di mana ia berbelok ke depan di bawah septum. Di sini bundel terbagi menjadi fasikuli kanan dan kiri, yang mengangkangi septum otot dan mengalir ke bawah di ventrikel kanan dan kiri, satu di setiap sisi septum. Akhirnya fasciculus mencapai otot papillary dari ventrikel dan membentuk pleksus serat Purkinje.
Fasikulus kanan memasuki ventrikel melalui trabekula septomarginal (pita moderator ), sedangkan yang kiri mencapai sisi kiri septum ventrikel dengan menembus bagian membran septum. Selubung fibrosa tipis melindungi nodus AV, bundel AV, dan serat Purkinje dari miokardium yang berdekatan. Karena bundel AV menegosiasikan urutan ritmis normal kontraksi atrium dan ventrikel, urutan ini mengalami gangguan besar ketika dirusak oleh penyakit.
Meskipun otot jantung membentuk syncytium yang jelas, otot atrium dan ventrikel terpisah karena adanya kerangka fibrosa jantung. Sistem konduktor adalah satu-satunya hubungan otot mereka. Sistem konduktor terjadi pada burung dan mamalia, karena mereka adalah hewan berdarah panas dan homoiotermik.
Kerangka Berserat Jantung (Gbr. 7-25):
Ini adalah kerangka fibrosa jantung tempat melekatnya otot-otot atrium dan ventrikel. Kerangka terdiri dari empat cincin berserat di dasar kedua ventrikel di sekitar lubang atrio-ventrikel paru, aorta, kanan dan kiri. Septum antero-posterior yang dikenal sebagai tendon infundibulum menghubungkan cincin paru dan aorta. Massa padat jaringan fibrosa yang dikenal sebagai trigonum fibrosum dextrum memanjang antara cincin aorta di depan dan cincin atrio-ventrikular kanan dan kiri di belakang. Bagian kiri jaringan ini dikenal sebagai trigonum fibrosum sinistrum. Pada beberapa hewan, seperti domba, tulang yang disebut os cordis muncul di trig onum fibrosum.

Tendon infundibulum dan trigonum fibrosum berlanjut di bawah dengan bagian membran septum ventrikel, dan di atas dengan septum inter-atrium.
Susunan Otot Jantung (Gbr. 7- 26):
Otot-otot atrium dan ventrikel disusun menjadi set fungsional yang berbeda.
Otot atrium:
Serat otot terdiri dari lapisan superfisial dan dalam.
Serabut superfisial berarah melintang , menutupi bagian depan dan belakang kedua atrium dan beberapa di antaranya masuk ke dalam septum inter-atrium. Kontraksi serabut-serabut ini mengurangi diameter transversal dan membentuk aksi sinkron kedua atrium.
Lapisan dalam terbatas pada setiap atrium dan disusun menjadi serat melingkar dan annular. Serat melingkar berbentuk ‘U’ terbalik dan ujungnya melekat pada cincin atrio-ventrikular. Setiap loop menutupi atap, dinding anterior dan posterior atrium. Kontraksi serat-serat ini mengurangi diameter vertikal atrium, dan membantu mendorong darah ke dalam ventrikel yang sesuai. Serat annular mengelilingi lubang vena, dan mencegah refluks darah ke dalam vena selama kontraksi sistolik atrium.
Otot ventrikel:
Serabut otot rumit dalam susunannya , dan berbagai penjelasan telah diberikan oleh pekerja yang berbeda.
Menurut otot ventrikel Leibel terdiri dari tiga lapisan-superfisial, tengah dan dalam. Lapisan superfisial dan dalam terus dekat dengan puncak jantung, setelah mereka mengalami jalur spiral. Lebih baik mempertimbangkan lapisan tengah terlebih dahulu.
Lapisan Tengah:
Ini adalah lapisan paling tebal dari ventrikel kiri, dan membentuk sebuah silinder yang menghadirkan margin bebas yang lebih rendah. Pada batas ini serabut-serabut otot menggelinding dengan sendirinya. Serat-serat lapisan tengah kiri timbul dari cincin atrio-ventrikel kiri, menyapu ke depan dan ke kanan di depan ventrikel kiri, dan dimasukkan di sekitar cincin paru dan aorta, dan tendon infundibulum. Bertindak dari cincin paru, kontraksi lapisan tengah cenderung menarik ventrikel kiri ke depan dan ke kanan. Ini menjelaskan mengapa jantung berputar ke depan dan ke kanan selama sistolik dan mendorong ke dinding dada.

Lapisan tengah ventrikel kanan tidak setebal sisi kiri. Serabut muncul dari cincin atrioventrikular kiri, dan terbagi menjadi rangkaian serabut longitudinal dan sirkular di sulkus interventrikular posterior. Serat-serat lon gitudinal berjalan ke bawah di septum interventrikular. Serabut sirkular mengelilingi ventrikel kanan, dan di sulkus interventrikularis anterior bersatu dengan serabut septum. Di sini mereka bersambung dengan lapisan tengah ventrikel kiri.
Hasil keseluruhan dari kontraksi lapisan tengah adalah untuk mengurangi diameter transversal kedua ventrikel; pengurangan lebih jelas di ventrikel kiri.
Lapisan Superfisial:
Serat muncul dari kerangka jantung dan menjalani jalur spiral. Mula-mula serabut-serabut itu menyapu ke bawah dan ke kanan melintasi permukaan inferior jantung. Mereka memutari batas bawah dan melewati ke kiri melintasi permukaan sternokostal. Saat mencapai puncak jantung, serabut superfisial membentuk pusaran di sekitar batas bawah lapisan tengah silinder, dan berbelok ke atas sebagai serat lapisan dalam.
Lapisan Dalam:
Serabut lapisan dalam naik tegak lurus ke serat superfisial, dan melekat pada otot papiler kedua ventrikel. Akhirnya, mereka mendapatkan keterikatan pada kerangka jantung melalui chordae tendinae dan berbagai katup.
Kontraksi simultan dari lapisan superfisial dan dalam menghasilkan pemendekan longitudinal ventrikel, karena kedua lapisan berorientasi pada sudut kanan satu sama lain. Sekarang diketahui bahwa jantung tidak benar-benar kosong selama kontraksi ventrikel. Beberapa volume sisa darah tetap berada di ventrikel pada akhir ejeksi. Volume stroke jantung manusia yang beristirahat adalah sekitar 60 ml. sedangkan volume diastolik darah di setiap ventrikel adalah sekitar 130 ml.
Pola pengosongan ventrikel:
Ventrikel kiri-Ini sebagian besar berkontraksi dengan mengurangi diameter melintang, melalui sebagian besar serat silinder tengah. Relatif sedikit pemendekan diameter longitudinal.
Ventrikel kanan-Pengeluaran darah dilakukan dengan dua metode:
(i) Ruang dipersingkat secara longitudinal oleh kontraksi simultan dari lapisan superfisial dan dalam.
(ii) Ventrikel kanan pada penampang berbentuk semi-lunar, dengan dinding septum cembung yang tetap dan dinding anterior bebas. Diameter transversal dikurangi dengan ‘tindakan di bawah’, ketika dinding bebas ventrikel kanan bergerak menuju permukaan cembung septum.
Pasokan Vaskular Otot Jantung:
Otot jantung banyak disuplai dengan kapiler dari arteri koroner. Endomisium setiap serat dilengkapi dengan darah dan kapiler limfatik. Dalam hal ini otot kardiac berbeda dengan otot volunter, dimana endomisium hanya mengandung kapiler darah. satu cm persegi. otot jantung disuplai oleh kapiler sepanjang 11 meter. Saat lahir, satu kapiler darah memasok empat atau lima serat otot jantung, sedangkan pada orang dewasa satu kapiler memasok hanya satu serat otot jantung.
Pasokan Saraf Otot Jantung:
Otot jantung disuplai oleh divisi simpatis dan parasimpatis (vagal) dari sistem saraf otonom. Serabut saraf mencapai serabut otot melalui pleksus jantung dan koroner. Atrium dan sistem konduksi jantung dipersarafi oleh saraf simpatis dan parasimpatis. Otot ventrikel, bagaimanapun, hanya disuplai oleh saraf simpatis.
Semua cabang jantung vagus dan simpatis mengandung serat sensorik dan motorik, kecuali cabang jantung dari ganglion simpatis serviks superior yang hanya mengandung serat motorik post-ganglionik.
Serabut motorik simpatis meningkatkan denyut jantung, dan menghasilkan vaso-dilatasi arteri koroner. Serabut sensorik simpatik menyampaikan sensasi nyeri dari jantung.
Serabut motorik parasimpatis mengurangi denyut jantung dan serabut sensoriknya berkaitan dengan refleks visceral yang menekan aktivitas mobil.
Regenerasi dan Pertumbuhan:
Karena fungsinya yang sangat terspesialisasi, otot jantung tidak dapat beregenerasi melalui pembelahan mitosis . Pada hipertrofi jantung, peningkatan sebagian besar otot disebabkan sepenuhnya oleh peningkatan ukuran serat individu.



