Fotosintesis: Proses Fotosintesis Dijelaskan (Dengan Diagram)!
Fotosintesis pada dasarnya adalah satu-satunya mekanisme input energi di dunia kehidupan. Fotosintesis (foto-cahaya, sintesis-penyatuan) adalah proses anabolik pembuatan senyawa organik di dalam sel yang mengandung klorofil dari karbon dioksida dan air dengan bantuan sinar matahari sebagai sumber energi. Persamaan sederhana fotosintesis adalah sebagai berikut:
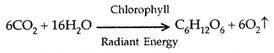 Sumber molekul oksigen adalah air dan bukan karbon dioksida seperti yang diyakini sebelumnya dibuktikan secara eksperimental pertama oleh Robert Hill (1937) dan kemudian dikonfirmasi oleh MD Kamen dan S. Ruben (1945), menggunakan teknik perunut di mana isotop berat oksigen 18 O digunakan. Tapi bukti eksperimental ini didasarkan pada saran karya CB Van Niel tentang fotosintesis bakteri (1930).
Sumber molekul oksigen adalah air dan bukan karbon dioksida seperti yang diyakini sebelumnya dibuktikan secara eksperimental pertama oleh Robert Hill (1937) dan kemudian dikonfirmasi oleh MD Kamen dan S. Ruben (1945), menggunakan teknik perunut di mana isotop berat oksigen 18 O digunakan. Tapi bukti eksperimental ini didasarkan pada saran karya CB Van Niel tentang fotosintesis bakteri (1930).
Ia mengemukakan bahwa pada tumbuhan hijau H 2 O merupakan sumber reduksi dan ketika dipecah menghasilkan (H) dan (OH), dan O 2 yang dikeluarkan oleh tumbuhan berasal dari air, bukan dari CO 2 . Pemisahan air dalam cahaya oleh tumbuhan hijau ini kemudian dikenal sebagai fotolisis air dan teorinya, teori fotolisis air Van Niel. 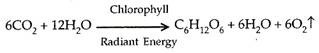
Kloroplas: Struktur dan Pigmen Fotosintetik:
Kloroplas adalah tempat pnotosintesis dan paling baik dicontohkan pada tumbuhan tingkat tinggi. Sebuah kloroplas ditutupi oleh selubung dua membran yang dipisahkan oleh ruang periplastidial 10-20 nm. Secara internal kloroplas mengandung matriks atau stroma yang tertanam sejumlah kantung membran pipih yang disebut tilakoid atau lamela. Permukaan luar tilakoid mengandung pigmen fotosintesis dan melayani ujung reaksi cahaya.
Stroma, di sisi lain, berkaitan dengan peristiwa reaksi gelap. Di daerah tertentu tilakoid ditumpuk membentuk grana. Tilakoid yang lebih panjang yang menghubungkan satu granum dengan yang lain meluas melalui stroma sehingga membran ini biasanya disebut sebagai tilakoid stroma.
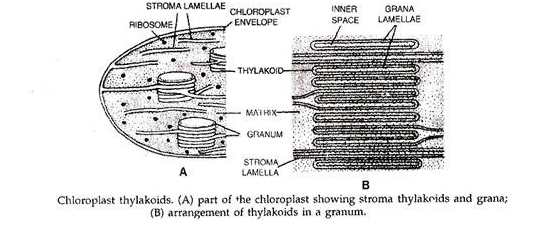
Pigmen Fotosintetik:
Pigmen fotosintesis hadir dalam membran tilakoid sebagian besar terdiri dari dua jenis klorofil hijau, Klorofil a (C 55 H 72 O 5 N 4 Mg) dan Klorofil b (C 55 H 70 O 6 N 4 Mg). Juga hadir pigmen kuning hingga oranye yang diklasifikasikan sebagai karotenoid.
Ada dua jenis karotenoid, karoten hidrokarbon murni dan xantofil yang mengandung oksigen. Karotenoid tertentu, terutama violaxanthin, sebuah xanthophyll, juga ada di selubung kloroplasi, memberikan warna kekuningan. Pada sebagian besar tumbuhan, termasuk ganggang hijau, β-karoten dan lutein adalah karotenoid yang paling melimpah di tilakoid.
Peristiwa Fotosintesis:
Fotosintesis terdiri dari dua jenis reaksi: yang bergantung pada cahaya dan yang tidak bergantung pada cahaya . Reaksi bergantung cahaya adalah reaksi fotokimia atau reaksi terang seperti yang kemudian disebut, yang berpuncak pada pembentukan NADPH 2 , ATP dan evolusi molekul oksigen.
NADPH 2 dan ATP kaya energi, setelah menangkap elektron yang tersedia saat cahaya mengenai klorofil. Mereka membentuk daya asimilasi, digunakan untuk fiksasi CO2 . Peristiwa fiksasi CO2 adalah reaksi bebas cahaya dan ditetapkan sebagai reaksi gelap.
Reaksi Ringan:
Reaksi terang terdiri dari dua fase: Fase I-Penyerapan energi (Penyerapan dan retensi cahaya oleh pigmen fotosintetik); dan Fase II-Transduksi energi (konversi energi cahaya yang diserap pada fase I menjadi energi kimia-ATP dan NADPH 2 melalui fotofosforilasi).
Fase I. Penyerapan energi:
Unit fotosintesis:
Peristiwa reaksi terang dimediasi melalui unit fotosintesis, unit fotosintesis menjadi kelompok molekul pigmen terkecil, bersama dengan zat asosiasi lipo-proteinnya, yang mampu menghasilkan aksi fotokimia (Photoact).
Istilah, tindakan fotokimia, berarti penyerapan dan migrasi kuantum cahaya oleh pusat perangkap, sebagai akibatnya elektron dilepaskan. Emerson dan Arnold berpikir bahwa unit fotosintesis (PSU) mengandung setidaknya 2500 molekul klorofil, tetapi penelitian terbaru oleh Bessel Kock menunjukkan bahwa unit fotosintesis hanya mengandung sekitar 250 molekul klorofil. Terjadinya PSU sebagai entitas morfologi yang berbeda diperoleh oleh Park dan rekan kerjanya dan mereka menamakannya quanta-some.
Penyerapan Cahaya oleh Pigmen:
Semua insiden cahaya pada permukaan fotosintesis tidak digunakan untuk fotosintesis. Sebagian besar hilang dan sebagian dipantulkan kembali. Sekali lagi, hanya sebagian kecil dari cahaya yang diserap digunakan untuk menggerakkan fotosintesis. Diperkirakan bahwa di bawah sinar matahari penuh, hanya sekitar 3% yang digunakan untuk keperluan fotosintesis.
Bergantung pada komposisi pigmen, berbagai kelompok tumbuhan menyerap dan memanfaatkan cahaya dari daerah spektral yang berbeda. Sebagian besar tumbuhan hijau menyerap cahaya dalam spektrum tampak (390-700 nm), sedangkan bakteri ungu menggunakan panjang gelombang mulai dari ultraviolet dekat hingga inframerah (800-950 nm).
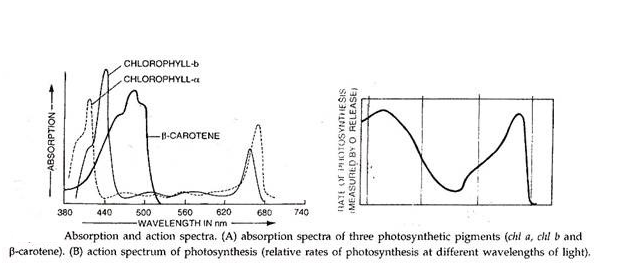
Kisaran spektrum di mana fotosintesis dapat terjadi disebut radiasi aktif fotosintesis. Tetapi seluruh rentang tidak dapat digunakan dalam fotosintesis. Tumbuhan hijau, misalnya, menyerap cahaya secara maksimal di daerah spektrum merah dan biru.
Sebuah studi tentang spektrum serapan menunjukkan hubungan kuantitatif antara panjang gelombang cahaya dan penyerapannya oleh pigmen yang bersangkutan. Jadi, kita melihat bahwa klorofil-a memiliki puncak serapan pada 660 nm dan 430 nm; klorofil-b pada 648 nm dan 456 nm; karoten pada 478 nm dan 449 nm dan xantofil sama dengan karoten.
Perangkap Cahaya:
Chlorophyll-fl memanfaatkan cahaya yang diserapnya sendiri dan juga cahaya yang ditransfer oleh pigmen lain. Terowongan cahaya dari pigmen lain ke klorofil a ini disebut perangkap cahaya atau wastafel cahaya. Perangkap cahaya menghasilkan efisiensi pemanenan cahaya yang jauh lebih baik, karena ini memastikan tunneling kuanta cahaya menuju satu molekul akseptor klorofil.
Reaksi terang sebenarnya terdiri dari dua reaksi fotokimia yang dipisahkan baik dalam ruang dan waktu. Mereka ditetapkan sebagai Photoact I dan Photoact II dan kedua reaksi tersebut dimediasi oleh dua sistem berbeda yang komposisinya berbeda dalam hal pigmen, pembawa elektron, dan mekanisme perangkap cahaya. Agen mediasi dari dua photoacts masing-masing disebut Fotosistem I dan Fotosistem II.
Fotosistem:
Konsep dua sistem foto berasal dari karya Emerson dan Lewis (1943). Bekerja pada spektrum aksi untuk pigmen Chlorella, mereka menemukan bahwa pada panjang gelombang cahaya antara 600 dan 680 nm (panjang gelombang yang sesuai dengan wilayah spektrum ‘merah’) evolusi oksigen mencapai maksimum.
Tetapi ketika cahaya dengan panjang gelombang di luar 680 nm, wilayah spektrum yang disebut ‘far-red’ disediakan; ada penurunan evolusi oksigen, yang menunjukkan berkurangnya efisiensi fotosintesis. Pengamatan ini telah diberi nama tetesan merah.
Kelompok penelitiannya menemukan bahwa jika cahaya dengan panjang gelombang yang lebih pendek diberikan pada waktu yang sama dengan panjang gelombang merah yang lebih panjang, fotosintesis bahkan lebih cepat daripada jika panjang gelombang itu sendiri disediakan. Sinergi atau peningkatan ini dikenal sebagai efek peningkatan Emerson. Dua pengamatan ini efek penurunan merah dan efek peningkatan menyebabkan indikasi pertama bahwa reaksi terang memiliki dua tempat aksi, satu di wilayah spektrum merah dan yang lainnya di daerah merah jauh.
Penjelasan yang ditawarkan untuk kedua efek ini adalah bahwa reaksi terang sebenarnya terdiri dari dua fotoakt, fotoakt I dan fotosistem II, diperantarai oleh dua fotosistem, fotosistem I dan fotosistem II.
Fotosistem I digerakkan oleh cahaya merah jauh dan ketika beroperasi sendiri, menghasilkan efek jatuh merah. Tetapi ketika beroperasi bersama dengan fotosistem II, yang berfungsi di wilayah merah, dihasilkan efek peningkatan. Pemisahan fisik kedua sistem foto telah berhasil dilakukan dan fungsinya diperjelas.
Fotosistem I terletak di membran tilakoid. Itu terdiri dari tiga bentuk klorofil-a, satu menyerap maksimal pada 683 nm, yang kedua menyerap maksimal pada 695 nm dan yang ketiga pada 670 nm. Yang terakhir disebut P-700. Fotosistem II terletak di stroma tilakoid. Itu terdiri dari dua bentuk klorofil-a dengan penyerapan maksimum pada 670 dan 690 nm.
Bentuk kedua diberi nama P-690. Setiap fotosistem memiliki tiga komponen: (i) pusat reaksi yang terbuat dari molekul klorofil khusus—dalam fotosistem I adalah molekul klorofil yang terikat protein, P-700; dalam fotosistem II adalah P-690.
Pusat reaksi adalah tempat aktual di mana energi cahaya diubah menjadi energi kimia, (ii) beberapa pembawa elektron—dalam fotosistem I, X, plastosianin, sitokrom-f dan ferrodoksin sebagai pembawa elektron; fotosistem II memiliki plastoquinone dan sitokrom b-559. (iii) klorofil dan karotenoid lain, yang hanya berfungsi untuk mentransfer cahaya yang diserapnya ke pusat aktif.
Fotosistem I mengambil bagian dalam fotofosforilasi siklik dan non-siklik. PS-I dapat melakukan fotofosforilasi siklik secara mandiri. Biasanya itu menggerakkan elektron dari fotosistem II ke NADP + .
Fotosistem II mengambil elektron yang dilepaskan selama fotolisis air. Hal yang sama diekstrusi pada penyerapan energi cahaya. Saat elektron yang diekstrusi melewati kompleks sitokrom, energi yang cukup dilepaskan untuk mengambil bagian dalam sintesis ATP dari ADP dan fosfat anorganik. Fotofosforilasi ini bersifat nonsiklik. PS II hanya dapat beroperasi bersamaan dengan PS I.
Fase II: Transduksi Energi:
Molekul tereksitasi dari P-700 dan P-690 mentransduser energinya untuk menghasilkan ATP dan NADPH 2 . Oksigen molekuler juga diproduksi tetapi keluar dari sistem fotosintesis. ATP dan NADPH 2 , bersama-sama membentuk daya asimilasi dan digunakan dalam fiksasi CO 2 dalam reaksi gelap .
Fotofosforilasi:
Fotofosforilasi adalah sintesis ATP yang digerakkan oleh cahaya atau energi cahaya. Ditemukan oleh Arnon et al pada tahun 1954. Fotofosforilasi terdiri dari dua jenis utama, siklik dan non-siklik.
Fotofosforilasi siklik:
Ini adalah proses fotofosforilasi di mana elektron yang dikeluarkan oleh pusat foto tereksitasi dikembalikan ke sana setelah melewati serangkaian pembawa elektron. Fotofosforilasi siklik hanya dilakukan oleh fotosistem I. Fotosentralnya P 700 mengeluarkan elektron dengan perolehan energi sebesar 23 kkal/mol setelah menyerap satu foton cahaya (hv).
Setelah kehilangan elektron, pusat foto menjadi teroksidasi. Elektron yang dikeluarkan melewati serangkaian pembawa termasuk X, ferredoxin, plasto quinone, kompleks sitokrom dan plastosianin sebelum kembali ke pusat foto.
Saat melewati antara ferredoxin dan plastoquinone dan / atau di atas kompleks sitokrom, elektron kehilangan energi yang cukup untuk membentuk ATP dari ADP dan fosfat anorganik.
Halobakteri atau bakteri halofil juga melakukan fotofosforilasi tetapi ATP yang dihasilkan tidak digunakan dalam sintesis makanan. Bakteri ini memiliki bacteriorhodopsin pigmen ungu yang melekat pada membran plasma. Saat cahaya jatuh pada pigmen, itu menciptakan pompa proton yang digunakan dalam sintesis ATP .
Fotofosforilasi Nonsiklik:
Ini adalah proses fotofosforilasi normal di mana elektron yang dikeluarkan oleh pusat foto yang tereksitasi tidak kembali ke sana. Fotofosforilasi non-siklik dilakukan dalam kolaborasi fotosistem I dan II. Elektron yang dilepaskan selama fotolisis air diambil oleh fotopusat PS II yang disebut P 600 .
Hal yang sama diekstrusi ketika pusat foto menyerap energi cahaya (hv). Elektron yang diekstrusi memiliki energi yang setara dengan 23 kkal/mol. Ia melewati serangkaian pembawa elektron—Q, PQ, kompleks sitokrom, dan plasma tosianin. Saat melewati kompleks sitokrom, elektron kehilangan energi yang cukup untuk sintesis ATP.
Elektron diserahkan ke photocentre P 700 dari PSI oleh plastocya nin. P 700 mengekstrusi elektron setelah menyerap energi cahaya. Elektron yang diekstrusi melewati X, pusat Fe-S A (feredoksin), dan NADP-reduktase yang menggabungkannya dengan NADP + . Huruf tersebut kemudian bergabung dengan H + (dilepaskan saat fotolisis) dengan bantuan NADP-reduktase membentuk NADPH.
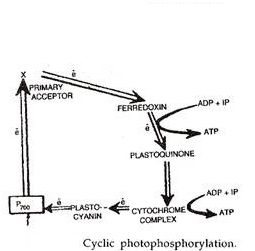
Sintesis ATP tidak langsung. Energi yang dilepaskan elektron sebenarnya digunakan untuk memompa ion H + melintasi membran tilakoid. Ini menciptakan gradien proton. Gradien memicu faktor kopling untuk mensintesis ATP dari ADP dan fosfat anorganik.
Evolusi Oksigen:
Oksigen yang dihasilkan selama fotosintesis berasal dari air dan merupakan bagian dari photoact II yang dimediasi oleh PSIL. Energi cahaya yang terperangkap oleh sistem ini menggairahkan P-690 dan dua elektron dikeluarkan. Energi tersebut digunakan untuk menghilangkan dua elektron dari hidrogen air dan mendorongnya ke tingkat energi yang lebih tinggi.
Pada titik ini, molekul oksigen yang lolos dari sistem fotosintesis terbentuk. Elektron melewati pembawa plastoquinone (PQ), sitokrom b-559, sitokrom-F, plastosianin dan akhirnya berakhir di P-700 untuk membawanya kembali ke keadaan dasar.
Jadi dalam fotosistem II, elektron yang membawa molekul klorofil tereksitasi ke keadaan dasar berasal dari fotolisis air. Aspek lain dari evolusi oksigen selama fotosintesis adalah hubungannya dengan keberadaan ion tertentu dalam medium seperti CI”, Mn 2+ dan bikarbonat.
2H 2 O———> O 2 + 4H + + 4e –
Kedua sistem foto yang bekerja bersama menghasilkan, untuk setiap dua putaran, dua molekul NADPH 2 , tiga ATP, dan satu molekul oksigen dari dua molekul air.
Reaksi Gelap atau Blackman:
Fase III: Stabilisasi Energi:
Produksi karbon dioksida menjadi karbohidrat adalah inti dari fase ini dan ini dicapai melalui penggunaan daya asimilasi (ATP dan NADPH 2 ) yang dihasilkan dalam reaksi terang. Stabilisasi energi ini adalah reaksi gelap.
Tidak memerlukan cahaya, melainkan daya asimilasi (ATP dan NADPH 2 ) yang dihasilkan selama fase fotokimia digunakan di sini dalam fiksasi dan reduksi CO 2 . Enzim yang diperlukan untuk proses tersebut terdapat dalam matriks atau stroma kloroplas. Ada dua jalur utama untuk biosintetik atau fase gelap—siklus Calvin dan siklus asam dikarboksilat C4 . Tumbuhan yang menunjukkan keduanya masing-masing disebut tumbuhan C 3 dan C 4 .
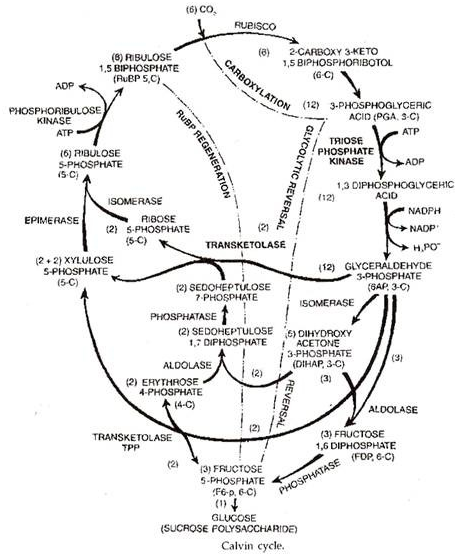
Siklus Calvin (siklus reduksi karbon fotosintesis atau jalur fotosintesis C 3 ).
Siklus ini ditemukan oleh Calvin, Benson dan rekannya menggunakan alga uniseluler Chlorella pyrenoidosa dan Scenedesmus obliques dan isotop radioaktif 14 C dengan waktu paruh lebih dari 5000 tahun.
Fase Siklus Calvin:
Siklus Calvin dibagi menjadi tiga fase berikut — karboksilasi, pembalikan glikolitik, dan regenerasi RuBP.
- Karboksilasi. Ini membutuhkan ribulosa-1, -bifosfat atau RuBP sebagai akseptor karbon dioksida dan RuBP karboksilase atau rubisco sebagai enzim. Enzim ini sebelumnya disebut karboksidismutase.
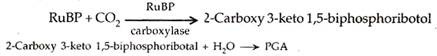
Karbon dioksida bergabung dengan ribulosa-1, 5-bifosfat untuk menghasilkan senyawa perantara sementara yang disebut 2-karboksi 3-keto 1,5-bifosforibotol. Perantara segera berpisah dengan adanya air untuk membentuk dua molekul asam 3-fosfogliserat atau PGA. Ini adalah produk fotosintesis stabil pertama.
- Pembalikan Glikolitik:
Proses yang terlibat dalam langkah atau fase ini adalah pembalikan proses yang ditemukan selama glikolisis bagian dari respirasi. Asam fosfogliserat atau PGA selanjutnya difosforilasi oleh ATP dengan bantuan enzim triose fosfat kinase. Ini menimbulkan asam 1,3-difosfogliserat.
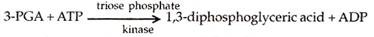
Asam difosfogliserat direduksi oleh NADPH melalui agen enzim triose fosfat dehidrogenase. Ini menghasilkan gliseraldehida 3-fosfat atau 3-fosfogliseraldehida.
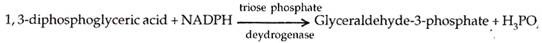
Gliseraldehida-3 fosfat adalah produk utama yang digunakan dalam sintesis karbohidrat dan lemak. Untuk membentuk karbohidrat, misalnya glukosa, sebagian diubah menjadi isomernya yang disebut dihidroksiaseton-3-fosfat. Enzim yang mengkatalisis reaksi adalah isomerase fosfat.

Kedua isomer memadat dengan adanya enzim aldolase membentuk fruktosa 1, 6-diphos phate .
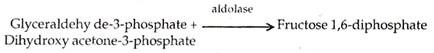
Fruktosa 1,6-difosfat (FDP) kehilangan satu gugus fosfat, membentuk fruktosa 6-fosfat (F 6-P) yang kemudian diubah menjadi glukosa-6-fosfat (G 6-P). Yang terakhir dapat menghasilkan glukosa atau menjadi bagian dari sukrosa dan polisakarida.

Karena glukosa adalah senyawa enam karbon, enam putaran siklus Calvin diperlukan untuk mensintesis satu molekulnya.
- Regenerasi RuBP:
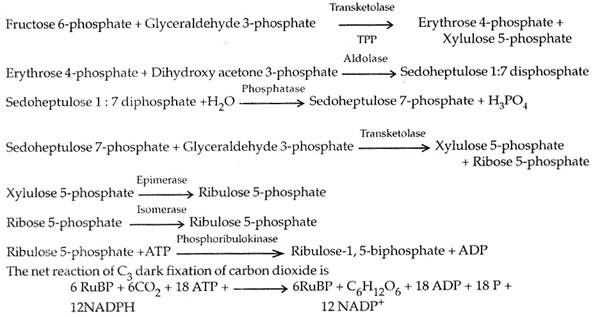
Fruktosa 6-fosfat (F 6-P) dan gliseraldehida 3-fosfat (GAP) bereaksi membentuk eritrosa 4-fosfat (E 4-P) dan xilulosa 5-fosfat (X 5-P). Eritrosa 4-fosfat bergabung dengan dihidroksi aseton 3-fosfat untuk menghasilkan sedoheptulosa 1 : 7-difosfat (SDP) yang kehilangan satu molekul fosfat dan menghasilkan sedoheptulosa 7-fosfat (S 7-P).
Sedoheptu kehilangan 7-fosfat bereaksi dengan gliseraldehida 3-fosfat menghasilkan xilulosa 5-fosfat (X 5-P) dan ribosa 5-fosfat. (R5-P). Keduanya diubah menjadi isomer ribulosa 5-fosfat (Ru 5-P). Ribulosa 5-fosfat mengambil fosfat kedua dari ATP untuk diubah menjadi ribulosa 1,5 bifosfat (RuBP).
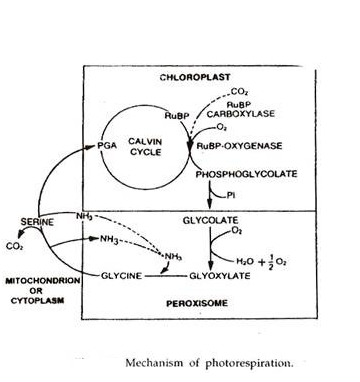
Fotorespirasi (Respirasi Terkait dengan Jaringan Fotosintetik):
Ditemukan oleh Decker dan Tio pada tahun 1959. Fotorespirasi adalah penggunaan oksigen dan pelepasan karbon dioksida yang bergantung pada cahaya oleh organ fotosintesis tanaman. Biasanya organ fotosintetik melakukan sebaliknya dalam cahaya yaitu, penyerapan CO 2 dan pelepasan O 2 .
Oleh karena itu, fotorespirasi sulit ditunjukkan. Hal ini disimpulkan dari (i) Penurunan laju fotosintesis bersih ketika konsentrasi oksigen meningkat dari 2—3% menjadi 21% (ii) Peningkatan evolusi CO 2 secara tiba-tiba ketika organ hijau terang dipindahkan ke gelap.
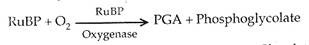
Tempat terjadinya fotorespirasi adalah kloroplas. Peroksisom diperlukan untuk menyelesaikan proses. RuBP karboksilase diubah menjadi RuBP oksigenase. Ini terjadi pada suhu tinggi dan konsentrasi oksigen tinggi. Pada suhu tinggi dan konsentrasi oksigen tinggi, afinitas RuBP karboksilase untuk CCX menurun dan afinitas terhadap O 2 meningkat.
Suhu tinggi terjadi di daerah tropis. Oleh karena itu, tanaman tropis adalah penderita utama. Pada suhu tinggi, RuBP karboksilase berfungsi sebagai oksigenase dan bukannya memperbaiki karbon dioksida, mengoksidasi ribulosa 1, 5-bifosfat untuk menghasilkan asam fosfogliserat dan fosfoglikolat.
Fosfoglikolat dihidrolisis menjadi glikolat. Glikolat biasanya masuk ke peroksisom sel mesofil dan membentuk glioksilat. Glioksilat diaminasi dan menimbulkan asam amino glisin. Di dalam mitokondria dan bahkan sitoplasma, dua molekul glisin memadat membentuk molekul serin, CO 2 dan amonia dilepaskan dalam proses tersebut. Serine selanjutnya dapat dideaminasi untuk membentuk PGA.
Yang terakhir masuk ke kloroplas untuk sintesis produk fotosintesis serta pembajakan fotores. Karena fotorespirasi melibatkan sintesis senyawa dua karbon, ini juga disebut siklus C 2 .
Fotorespirasi tidak menghasilkan energi atau mengurangi daya. Sebaliknya, itu menghabiskan energi. Selanjutnya, itu membatalkan pekerjaan fotosintesis. Ini dapat mengurangi fotosintesis hingga 50%. Oleh karena itu, fotorespirasi adalah proses yang sangat boros.
Ini hanya terjadi pada tanaman C 3 . Tumbuhan C 4 telah mengatasi masalah fotorespirasi dengan melakukan Siklus Calvin di bagian dalam daun (sel seludang bundel) di mana suhu dan oksigen lebih rendah. Mereka selanjutnya memastikan pasokan CO2 yang tinggi ke sel -sel yang melakukan siklus Calvin.
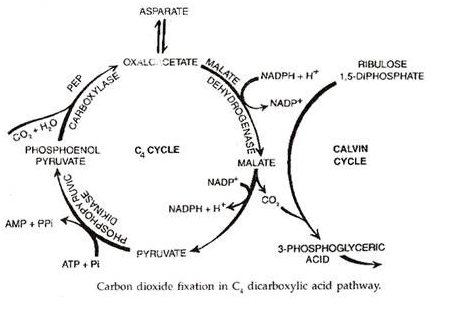
C 4 -Jalur Asam Dikarboksilat:
(Jalur Penetas Slack, Jalur C 4 )
Itu dikerjakan oleh Hatch and Slack (1965,1967). Kortschak et al (1965) menemukan bahwa karbon dioksida berlabel ( 14 C0 2 ) diasimilasi oleh daun Tebu pertama kali muncul dalam senyawa 4-karbon asam oksalo-asetat (OAA atau oksaloasetat).
Hatch dan Slack menemukan cara reguler fiksasi CO 2 di sejumlah tanaman tropis, baik monokotil maupun dikotil, misalnya Jagung, Tebu, Sorgum, Panikum, Pentiisetum, Atriplex, Amaranthus, Salsola dll. Tumbuhan ini adalah disebut tanaman C 4 karena produk fotosintesis stabil pertama adalah senyawa 4-karbon. Tumbuhan lainnya adalah tumbuhan C 3 . Tumbuhan C 4 sering hidup di habitat yang panas, gersang, dan asin. Mereka memiliki anatomi Kranz.
Dalam anatomi Kranz, mesofil tidak berdiferensiasi dan sel-selnya muncul di lapisan konsentris di sekitar bundel vaskular yang memiliki sel selubung bundel besar. Sel mesofil dan seludang berkas dihubungkan oleh plasmodesmata atau jembatan sitoplasma.
Kloroplas sel mesofil lebih kecil. Mereka memiliki grana yang berkembang dengan baik dan retikulum perifer tetapi tidak memiliki pati. Kloroplas sel selubung bundel lebih besar. Mereka memiliki grana yang tidak jelas, retikulum perifer, dan butiran pati.
Pada tanaman C4, fiksasi awal karbon dioksida terjadi pada sel mesofil . Akseptor utama CO 2 adalah fosfoenol piruvat atau PEP. Ini bergabung dengan karbon dioksida di hadapan karboksilase PEP atau pepco untuk membentuk asam oksalo-asetat atau oksaloasetat.
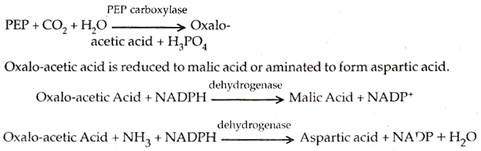
Asam malat atau asam aspartat ditranslokasi ke sel seludang bundel melalui plasmodesmata. Di samping sel-sel selubung bundel mereka mengalami dekarboksilasi (dan dideaminasi dalam kasus asam aspartat) untuk membentuk piruvat dan CO 2 .
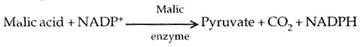
CO 2 lagi tetap di dalam sel selubung bundel melalui siklus Calvin. RuBP siklus Calvin disebut akseptor sekunder atau akhir CO 2 pada tanaman C 4 . Piruvat dikirim kembali ke sel mesofil. Di sini, diubah menjadi fosfoenol piruvat. Energi diperlukan untuk ini. Hal yang sama disediakan oleh ATP. Yang terakhir diubah menjadi AMP (adenosine monophosphate).
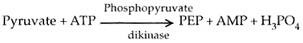
Konversi AMP menjadi ATP membutuhkan energi dua kali lipat daripada energi ADP menjadi ATP. Oleh karena itu, kebutuhan energi sebenarnya sama dengan dua molekul ATP. Energi ini merupakan tambahan dari 3 ATP yang diperlukan untuk fiksasi satu molekul CO 3 melalui siklus Calvin. Oleh karena itu, tanaman C 4 mengkonsumsi 5 molekul ATP per molekul CO 2 tetap, bukan 3 molekul ATP untuk tanaman C 3 . Untuk pembentukan molekul glukosa, tanaman C 4 membutuhkan 30 ATP sedangkan tanaman C 3 hanya menggunakan 18 ATP.
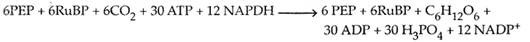
Pentingnya:
- Tumbuhan C 4 dianggap memiliki efisiensi fotosintesis yang lebih besar , karena mereka dapat memanfaatkan CO, sampai kadar 5 ppm tercapai tetapi tumbuhan siklus Calvin tidak dapat memanfaatkan C0 2 jika kadarnya turun di bawah 40-50 ppm.
- C 4 -tanaman dapat memanfaatkan intensitas cahaya yang lebih besar dan suhu optima untuk fotosintesis melebihi tanaman C 3 .
- Kloroplas tanaman ini tampaknya menghasilkan lebih banyak ATP yang tentu saja meningkatkan kerja seluler.
- Kehadiran retikulum perifer yang luas dalam kloroplas tanaman ini secara tidak langsung menunjukkan pengangkutan produk yang lebih cepat dan oleh karena itu penggunaan cahaya dan CO2 yang lebih besar .
Metabolisme Asam Crassulacean:
Jalur ini yang dikembangkan oleh Ranson dan Thomas (1960) dan Rouham, Vines and Black (1973) ditemukan pada sukulen, kebanyakan anggota Crassulaceae (Bryophylnm dan Seditm) dan beberapa anggota Bromeliaceae, seperti nanas. Ini adalah perangkat yang dirancang untuk memenuhi tekanan transpirasi berat , yang timbul dari lingkungan xerophytic mereka.
Tumbuhan ini memperoleh kebutuhan CO 2 pada malam hari ketika stomata mereka tetap terbuka dan saat penumpukan CO 2 terjadi, getah sel berubah menjadi asam. Proses ini disebut pengasaman gelap. Pada siang hari berikutnya, stomata tetap tertutup, meminimalkan kerugian transpirasional, tetapi dengan adanya cahaya, CO2 yang diserap selama malam sebelumnya digunakan untuk keperluan fotosintesis, proses deasidifikasi cahaya kemudian terjadi.
Dengan demikian, ada jeda waktu antara penyerapan dan reduksi CO2 . Pengaturan ini membantu mengurangi stres transpirasi tetapi bertanggung jawab atas pertumbuhan yang sangat lambat dari tanaman ini. Di bawah ini diberikan penjelasan singkat tentang jalur ini.
Fase I: Pengasaman gelap:
Pada fase ini, pati cadangan dipecah menjadi fosfoenoplyruvat (PEP) melalui sejumlah reaksi pernapasan. PEP menerima CO2 atmosfer yang diserap oleh tanaman dan menghasilkan asam malat. Ini terjadi dalam kegelapan ketika stomata terbuka.
Pati———– > PEP.
PEP + NADPH 2 + CO 2 dari atmosfer –> Asam malat + NADP.
Fase II: Deasidifikasi ringan:
Selama fase ini, yang terjadi pada cahaya saat stomata tertutup, asam malat mengalami dekarboksilasi menjadi asam piruvat dan CO 2 . CO 2 yang dilepaskan disalurkan melalui siklus Calvin untuk mensintesis heksosa. Asam piruvat digunakan untuk membangun pati yang stoknya habis lebih awal.
Asam malat –> Asam piruvat + NADPH 2 + CO 2 pada tanaman
Peristiwa siklus Calvin:
6 CO 2 + 12 NADPH 2 + 18 ATP + 11 H 2 O Fruktosa -6-fosfat + 12 NADP + 18 ADP + 17 Pi
Daya asimilasi yang dibutuhkan untuk siklus Calvin disediakan oleh peristiwa deasidifikasi ringan.
Prinsip atau Hukum Faktor Pembatas:
Nilai optimal suatu faktor tidak pernah konstan. Itu tergantung pada besarnya faktor lain. Kami dapat terus meningkatkan besarnya satu atau lebih faktor tanpa mempengaruhi laju reaksi. Dalam kasus seperti itu ditemukan bahwa faktor yang disebut faktor pembatas memegang keseimbangan.
Faktor pembatas didefinisikan sebagai faktor yang kekurangan sedemikian rupa sehingga peningkatan besarnya secara langsung meningkatkan laju proses. Pengaruh faktor pembatas dipelajari oleh Blackmann pada tahun 1905. Dia merumuskan prinsip faktor pembatas yang menyatakan bahwa ketika suatu proses dikondisikan untuk kecepatannya oleh sejumlah faktor terpisah, laju proses dibatasi oleh kecepatan proses. faktor paling lambat. Dengan kata lain laju proses fisiologis dibatasi pada waktu tertentu oleh satu dan hanya satu faktor yang kurang.



