Jalur dan Mekanisme Translokasi Floem pada Tumbuhan!
Nutrisi organik yang paling umum ditranslokasi pada tanaman adalah sukrosa. Saluran transportasi adalah tabung saringan (pada tanaman berbunga) dan sel saringan (pada tanaman vaskular tidak berbunga) floem. Hal itu dibuktikan pertama kali oleh Czapek (1897).
Beberapa teori telah dikemukakan untuk menjelaskan mekanisme translokasi nutrisi organik melalui floem misalnya, difusi, difusi aktif, aliran protoplasma, aliran antarmuka, elektrosmosis, untaian transelular, protein kontraktil, aliran massa. Yang penting adalah sebagai berikut:
1. Hipotesis Aliran Sitoplasma:
Itu dikemukakan oleh de Vries (1885) dan kemudian dikembangkan oleh Curtis (1929-35). Hipotesis menganggap transportasi berlangsung dengan kombinasi dua kekuatan, difusi dan aliran sitoplasma. Aliran sitoplasma membawa larutan organik atau bahan makanan dari satu ujung ke ujung lain dari sel tabung saringan dari mana mereka berdifusi sesuai dengan gradien konsentrasinya ke sel tabung saringan yang berdekatan melalui pelat saringan.
Hipotesis dapat menjelaskan pergerakan dua arah zat dalam elemen saringan yang sama. Kelemahan penting dari teori ini adalah (i) Tingkat aliran sitoplasma terlalu lambat untuk memperhitungkan transportasi floem (5 cm/jam dibandingkan dengan 50-150 cm/jam transportasi floem), (ii) Meskipun Thaine (1954) telah mengamati aliran sitoplasma terjadi pada beberapa tabung ayakan yang matang, biasanya tidak ada di dalamnya, (iii) Eksudat floem tidak mengandung sitoplasma.
2. Hipotesis Streaming Transelular:
Hipotesis telah dikemukakan oleh Thaine (1962, 1969). Ia percaya bahwa tabung saringan memiliki untaian tubular yang terus menerus dari satu sel tabung ke sel lainnya melalui pori-pori saringan. Untaian tubular atau transelular menunjukkan semacam gerakan peristaltik yang membantu lewatnya zat organik. Untaian transeluler, bagaimanapun, tidak diamati dalam tabung saringan.
3. Hipotesis Aliran Massa atau Aliran Tekanan:
Itu dikemukakan oleh Munch (1927,1930). Menurut hipotesis ini, zat organik berpindah dari daerah tekanan osmotik tinggi ke daerah tekanan osmotik rendah dalam aliran massa karena perkembangan gradien tekanan turgor. Hal ini dapat dibuktikan dengan mengambil dua osmometer yang saling berhubungan, satu dengan konsentrasi zat terlarut tinggi dan yang lainnya dengan konsentrasi osmotik kecil.
Dua osmometer alat ditempatkan dalam air. Lebih banyak air memasuki osmometer yang memiliki konsentrasi zat terlarut tinggi dibandingkan dengan yang lain. Oleh karena itu, ia akan memiliki tekanan turgor tinggi yang memaksa larutan untuk masuk ke osmometer kedua melalui aliran massa. Jika zat terlarut diisi ulang dalam osmometer donor dan diimobilisasi dalam osmometer penerima, aliran massa dapat dipertahankan tanpa batas.
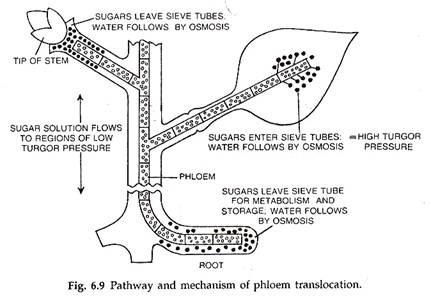
Sistem tabung saringan sepenuhnya disesuaikan dengan aliran massa zat terlarut. Di sini vakuola sepenuhnya permeabel karena tidak adanya tonoplas (Esau, 1966). Konsentrasi osmotik tinggi yang terus-menerus ada di sumber atau wilayah pasokan, misalnya sel mesofil (karena fotosintesis).
Zat organik yang ada di dalamnya dilewatkan ke tabung saringan melalui sel pendampingnya melalui proses aktif. Oleh karena itu, konsentrasi osmotik yang tinggi berkembang di tabung saringan sumber. Tabung saringan menyerap air dari xilem sekitarnya dan mengembangkan tekanan turgor yang tinggi.
Hal ini menyebabkan aliran larutan organik menuju daerah bertekanan turgor rendah. Tekanan turgor yang rendah dipertahankan di daerah sink dengan mengubah zat organik yang larut menjadi bentuk yang tidak larut. Air melewati kembali ke xilem.
Bukti:
(i) Tabung saringan mengandung zat terlarut organik di bawah tekanan karena cedera menyebabkan eksudasi larutan yang kaya zat terlarut organik.
(ii) Arah aliran zat terlarut organik selalu menuju gradien konsentrasi. Penurunan konsentrasi 20% diamati oleh Zimmermartn (1957) pada jarak delapan meter.
(iii) Defoliasi pucuk menyebabkan hilangnya gradien konsentrasi pada floemnya.
(iv) Bennet (1937) mengamati virus bergerak dalam floem dalam aliran massa searah dengan pergerakan zat terlarut organik dengan kecepatan sekitar 60 cm/jam.
(v) Semua zat yang terlarut dalam tabung ayakan diketahui bergerak dengan kecepatan yang sama dengan sedikit perbedaan.
(vi) Hipotesis dapat disimulasikan secara eksperimental.
Keberatan:
(i) Vakuola sel tabung saringan yang berdekatan tidak kontinu. Sitoplasma yang ada di dekat pelat saringan memberikan perlawanan terhadap aliran massa.
(ii) Catalado et al (1972) telah mengamati bahwa laju aliran air (72 cm/jam) dan zat terlarut (35 cm/jam) berbeda dalam tabung saringan yang sama.
(iii) Transpor floem tidak dipengaruhi oleh defisit air.
(iv) Sel-sel di ujung sumber aliran massa harus bombastis tetapi sel-sel tersebut sering ditemukan lembek dalam kasus umbi, umbi, dll yang berkecambah.



