Mitokondria: Distribusi, Morfologi, Fungsi dan Asal Mitokondria!
Kоlliker (1880) adalah orang pertama yang mengamati butiran (mitokondria) pada sel otot serangga. Flemming (1882) menamakan mitokondria sebagai fila. Altmann pada tahun 1894 mengamati mereka dan mereka disebut bioplast butiran Altmann.
Istilah mitokondria diterapkan oleh Benda (1897-98) pada granula ini yang digambarkan sebagai sitomikrosom oleh Velette St. George. Benda mewarnai mitokondria dengan alizarin dan kristal violet. Kingsbury (1912) menghubungkannya dengan respirasi seluler dan Warburg (1913) mengamati keberadaan enzim respirasi. Pada tahun 1934 Bensley dan Horr mengisolasi mitokondria dari sel hati dan Porter dan Palade menggambarkan struktur mikroskop elektron mereka.
Distribusi:
Biasanya mitokondria tersebar merata di sitoplasma. Namun, mereka mungkin terlokalisasi di wilayah tertentu. Dalam tubulus kontortus proksimal ginjal, mereka ditemukan di daerah basal sel, di seberang kapiler ginjal. Pada otot rangka mereka terletak di antara miofibril. Pada otot terbang serangga beberapa mitokondria besar bersentuhan dengan masing-masing fibril.
Pada otot jantung, mitokondria terletak di celah antara miofibril, banyak tetesan lipid yang berhubungan dengan mitokondria. Pada banyak sperma, mitokondria melebur menjadi satu atau dua struktur yang terletak di bagian tengah sperma, mengelilingi filamen ketiak. Dalam sel kolumnar atau prismatik, mereka diorientasikan sejajar dengan sumbu panjang sel. Dalam leukosit mereka tersusun secara radial.
Orientasi:
Mitokondria mungkin memiliki orientasi yang kurang lebih pasti. Misalnya dalam sel silinder umumnya berorientasi pada arah basal-apikal, sejajar dengan sumbu utama. Dalam leukosit, mitokondria tersusun secara radial sehubungan dengan sentriol. Telah dikemukakan bahwa orientasi ini bergantung pada arah arus difusi di dalam sel dan terkait dengan organisasi submikroskopik dari matriks sitoplasma dan sistem vakuolar.
Plastisitas Mitokondria dalam Sel:
Lewis dan Lewis (1914-15) menyimpulkan bahwa mitokondria adalah benda yang sangat bervariasi, yang terus bergerak dan berubah bentuk dalam sitoplasma. Tidak ada jenis mitokondria yang pasti, karena salah satu jenis dapat berubah menjadi jenis lainnya. Mereka tampaknya muncul di sitoplasma dan digunakan oleh aktivitas seluler.
Bentuknya bisa berubah lima belas sampai dua puluh kali dalam sepuluh menit; itu bisa dengan kalium permanganat dan perubahan osmotik. Frederic (1958), Littre (1954), Tobioka dan Biesels (1956) mempelajari efek sejumlah besar bahan kimia dan fisika terhadap perilaku mitokondria. Beberapa bahan, seperti deterjen menunjukkan beberapa efek in vivo seperti pada mitokondria yang diisolasi dari homogenat.
Morfologi:
Membentuk:
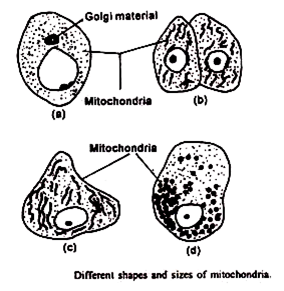
Bentuknya bervariasi tetapi khas untuk sel atau jenis jaringan, ini juga bergantung pada lingkungan atau kondisi fisiologis. Secara umum mereka berserabut atau granular. Mereka mungkin membengkak di salah satu ujungnya menjadi berbentuk tongkat atau berlubang di salah satu ujungnya untuk mengambil bentuk raket tenis. Mereka mungkin menjadi vesikular dengan munculnya zona bening sentral. Mitokondria berbentuk batang juga dapat diamati.
Ukuran:
Ukuran mitokondria juga bervariasi. Pada sebagian besar sel, lebarnya relatif konstan, sekitar 0,5µ, tetapi panjangnya bervariasi dan terkadang mencapai maksimum 7µ. Ukuran sel juga tergantung pada tahap fungsional sel. Mitokondria yang sangat tipis, sekitar 0,2µ, atau batang tebal 2µ juga terlihat.
Ukuran dan bentuk mitokondria tetap ditentukan oleh tekanan osmotik dan pH fiksatif. Dalam asam, mitokondria terfragmentasi dan menjadi vesikular. Panjang mitokondria di hati tikus biasanya 3,3µ; pada pankreas eksokrin mamalia, panjangnya sekitar 10µ dan pada oosit Amphibis, panjangnya sekitar 20 hingga 40µ.
Nomor:
Mitokondria ditemukan dalam sitoplasma semua sel yang berespirasi secara aerobik, kecuali bakteri yang enzim-enzim pernapasannya terletak di membran plasma. Kandungan mitokondria sel sulit ditentukan, tetapi secara umum bervariasi dengan jenis sel dan tahap fungsional.
Diperkirakan bahwa di hati mitokondria merupakan 30 sampai 35 persen dari total isi sel dan di ginjal, 20 persen. Dalam jaringan limfoid nilainya jauh lebih rendah. Dalam homogenat hati tikus terdapat sekitar 8,7×10’° mitokondria per gram jaringan segar. Sel hati yang normal mengandung sekitar 100 hingga 1600 mitokondria, tetapi jumlah ini berkurang selama regenerasi dan juga pada jaringan kanker.
Pengamatan terakhir ini mungkin terkait dengan penurunan oksidasi yang menyertai peningkatan glikolisis anaerobik pada kanker. Temuan menarik lainnya adalah adanya peningkatan jumlah mitokondria di otot setelah pemberian berulang hormon tiroid, tiroksin. Peningkatan jumlah mitokondria juga ditemukan pada hipertiroidisme manusia.
Jadi sel dengan aktivitas metabolisme tinggi memiliki jumlah mitokondria yang tinggi, sedangkan sel dengan aktivitas metabolisme rendah memiliki jumlah mitokondria yang lebih rendah. Telur bulu babi besar memiliki 13.000-14.000, sedangkan tubulus ginjal memiliki 300-400. Di dalam sperma terdapat sedikitnya 20-24 mitokondria sementara di beberapa oosit terdapat sekitar 300.000. Dalam kekacauan kekacauan protozoa ada sekitar 500.000 mitokondria. Beberapa sel alga hanya mengandung satu mitokondria.
Struktur Mitokondria:
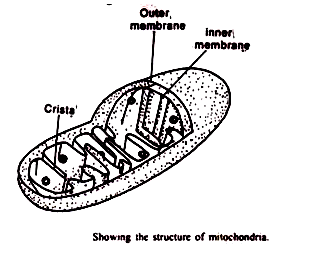
Sebuah mitokondria khas dalam sosis berbentuk dengan diameter rata-rata sekitar 0,5 (i. Ketika itu benar tetap dalam cairan yang mengandung osmium dan dipelajari di bawah mikroskop elektron yang mengungkapkan bahwa hampir tidak ada perbedaan antara mitokondria tumbuhan dan hewan. Dalam kedua kasus mitokondria dibatasi oleh dua membran yaitu membran luar dan membran dalam.
Ruang antara dua membran disebut ruang luar atau ruang antar membran. Itu diisi dengan cairan encer, dan lebarnya 40-70A 0 . Ruang yang dibatasi oleh membran dalam disebut ruang dalam atau ruang membran dalam.
Ruang membran bagian dalam diisi dengan matriks yang mengandung butiran padat (300-500A 0 ), ribosom dan DNA mitokondria. Butiran terdiri dari garam anorganik yang tidak larut dan diyakini sebagai tempat pengikatan ion divalen seperti Mg ++ dan Ca ++ .
 Dalam beberapa kasus, ternyata mengandung polimer gula. Sisi membran dalam yang menghadap sisi matriks disebut sisi-M, sedangkan sisi yang menghadap ruang luar disebut sisi-C mati. Dua hingga enam molekul DNA sirkular telah diidentifikasi dengan mitokondria. Cincin ini mungkin terbuka atau dalam konfigurasi bengkok. Mereka mungkin hadir bebas dalam matriks atau mungkin melekat pada membran. Enzim dari siklus Krebs terletak di matriks.
Dalam beberapa kasus, ternyata mengandung polimer gula. Sisi membran dalam yang menghadap sisi matriks disebut sisi-M, sedangkan sisi yang menghadap ruang luar disebut sisi-C mati. Dua hingga enam molekul DNA sirkular telah diidentifikasi dengan mitokondria. Cincin ini mungkin terbuka atau dalam konfigurasi bengkok. Mereka mungkin hadir bebas dalam matriks atau mungkin melekat pada membran. Enzim dari siklus Krebs terletak di matriks.
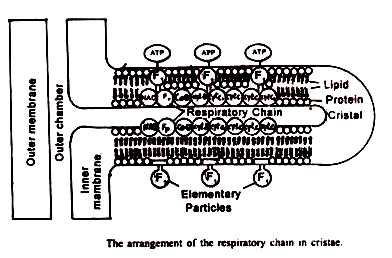
Membran bagian dalam terlempar ke atas menjadi serangkaian lipatan, yang disebut krista mitokondria, yang menonjol ke dalam ruang dalam. Rongga krista disebut ruang antar krista, dan bersambung dengan ruang antar membran.
Ruang dan susunan puncak bervariasi dan mungkin dari jenis berikut:
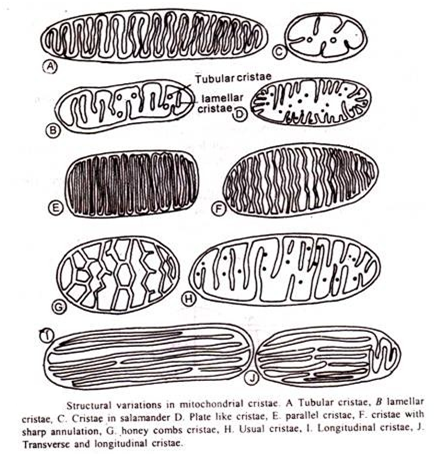
(i) Sejajar dengan sumbu panjang mitokondria seperti pada neuron dan sel otot lurik.
(ii) Tersusun secara konsentris seperti pada matriks spermatid tertentu.
(iii) Terjalin membentuk vili seperti pada Amoeba.
(iv) Cristae berupa vesikel yang membentuk jaringan ruang-ruang yang saling berhubungan seperti pada sel-sel kelenjar paratiroid dan W.Ð’. C. manusia.
(v) Tersusun secara tubular tetapi tegak lurus terhadap sumbu mitokondria seperti pada sel-sel kelenjar adrenal.
(vi) Didistribusikan secara acak seperti pada sel ginjal serangga dan sel hati.
(vii) Cristae sangat kecil dan tidak beraturan seperti pada sel interstitial Opossum.
(viii) Jarang dinding mitokondria mulus tanpa krista. Jumlah dan ukuran krista dalam mitokondria secara langsung mempengaruhi efisiensi. Semakin besar dan besar krista, semakin cepat kecepatan reaksi oksidasi.
(ix) Tegak lurus terhadap sumbu panjang mitokondria.
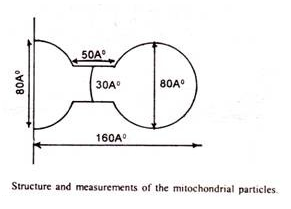
Partikel mitokondria:
Menurut deskripsi (Green dan Perdue, 1966) permukaan luar membran luar dan permukaan dalam membran dalam seharusnya ditutupi dengan ribuan partikel kecil. Mereka yang berada di membran luar digambarkan sebagai tangkai yang lebih sedikit dan disebut subunit Parson.
Mungkin ada sebanyak 10.000 sampai 100.000 partikel per mitokondria. Namun, penelitian terbaru menunjukkan bahwa partikel tanpa tangkai tidak ada. Partikel membran dalam bertangkai disebut subunit Femandez-Moran, Partikel elementer, partikel FI atau oksiosom atau ETP atau partikel transpor elektron. Partikel-partikel ini berdiameter sekitar 84A° dan berjarak teratur dengan interval 10 nm pada membran dalam. Mungkin ada sebanyak 10 4 sampai 10 5 partikel elementer per mitokondria.
Isolasi Mitokondria:
Mitokondria dapat diisolasi dari sel dalam bentuk hidup untuk studi fisiologisnya. Sel-sel pertama diperlakukan dengan deoxycholate untuk kerusakannya. Kemudian mereka dilewatkan dalam larutan sukrosa. Homogenat harus disentrifugasi selama 10 menit dengan kecepatan 6000 X g. Dari homogenat zat atas ini disentrifugasi dengan kecepatan 8500 X g selama 10 menit.
Setelah sentrifugasi ini, fraksi mikrosomal atas dibuang sedangkan fraksi bawah terdiri dari mitokondria dan partikel lain seperti lisosom. Fraksi ini dilewatkan melalui gradien sukrosa. Fraksi mitokondria kemudian disentrifugasi dengan kecepatan 10.000 X g selama 3 jam. Bagian atas bahan yang disentrifugasi ini memiliki mitokondria dan bagian bawah lisosom.
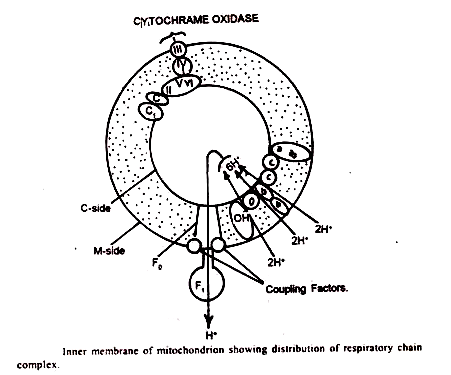
Kompleks Rantai Pernafasan:
Hijau et al. telah mengenali lima kompleks utama yang, jika dicampur dalam rasio yang tepat, dapat menyusun kembali untuk membentuk ETC.
Kompleks ini adalah:
- Kompleks 1 (NADH-Q-reduktase):
Ini adalah kompleks terbesar, dengan berat molekul sekitar 500.000 dan struktur terdiri dari 15 subunit. Ini berisi sebagai kelompok prostetik flavin mononukleotida (FMN) dan enam pusat besi-sulfur. Situs reaksi NADH terletak di sisi M mitokondria.
Kontak antara NADH dan CoQ tampaknya dibuat di tengah membran. Kompleks 1 menjangkau membran mitokondria bagian dalam dan mampu mentranslokasi proton melewatinya, dari sisi M ke sisi С mitokondria.
- Kompleks II (Suksinat-Q-reduktase):
Kompleks ini terdiri dari dua polipeptida dengan berat molekul 97000. Ini mengandung flavin adenine dinucleotide (FAD) dan tiga pusat besi-sulfur (Fe-SS1, Fe-SS2 dan Fe-SS3). Situs pengikatan suksinat terjadi di sisi M.
Ketiga pusat Fe-S berada di sisi M dan terdapat interaksi yang erat antara pusat Fe-S 3 dan CoQ. Berbeda dengan kompleks I, succnate-Q-reductase ternyata tidak mampu mentranslokasi proton melintasi membran.
- Kompleks III (Q H2-sitokrom-C-reduktase) :
Kompleks ini berisi sejumlah kiriman dengan berat molekul 280.000. Ini mengandung sitokrom b, sitokrom C, dan protein belerang besi. Ada dua jenis sitokrom b, yaitu but (transducing cytochrome b) dan bk (Keilin-type cytochrome b). Heme yang mengandung bagian sitokrom C,’ yang mentransfer elektron ke sitokrom – c, terletak di sisi С mitokondria.
- Kompleks IV (Silokrom-C-oksidase) :
Ini memiliki dua sitokrom, a dan 3 , dan dua atom tembaga. Berat molekulnya sekitar 200.000. Akan tetapi, a dan a 3 tidak pernah dipisahkan dan oleh karena itu keduanya harus dianggap sebagai bagian dari kompleks yang sama. Kompleks IV diperkirakan melintasi membran mitokondria, menonjol di kedua permukaan.
Dalam percobaan dengan mitokondria ragi telah ditunjukkan bahwa sitokrom-c-oksidase terbuat dari tujuh subunit. Tujuh subunit disusun membran dalam urutan fungsional, bersentuhan dengan sitokrom Ñ di sisi С. Elektron kemudian berpindah ke sitokrom a kemudian ke Cu ++ dan akhirnya ke sitokrom a., oksigen di sisi M.
- Kompleks V (kompleks ATPase):
Menuju sisi M, membran bagian dalam mengandung partikel bertangkai bulat- dikenal sebagai partikel F, atau partikel Fernandez-Moran. Setiap F, partikel terdiri dari kepala, tangkai dan alas. Telah ditunjukkan bahwa kompleks V identik dengan partikel F1 . Ini adalah empat faktor penyusun pada sisi M yang ditetapkan sebagai F 1 F 2 F 3 atau OSCP (protein pemberi sensitif oliligomisin) dan F 6 .
Topi baja, faktor kopling F 1 adalah ATPase yang tepat. F, berisi lima jenis subunit, a dengan berat molekul 53.000 dalton, β dengan berat molekul 50.000 dalton, y dengan berat molekul 33.000 delton, β dengan berat molekul 17.000 dalton dan e dengan berat molekul 7.000 delton. Selain subunit tersebut terdapat ATPase inhibitor (I) dengan berat molekul 10.000 dalton. Inhibitor ini dapat dihilangkan saat merawat dengan tripsin.
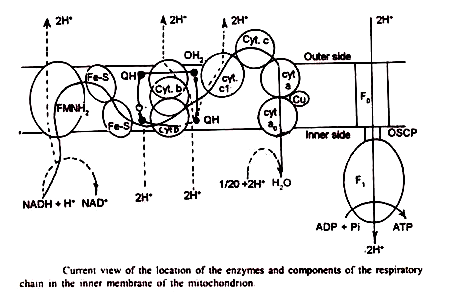
Tangkai yang terbuat dari protein menghubungkan kepala – potongan ke pangkal. Porsi ini sesuai dengan OSCP (oilgomisin –protein pemberi sensitif) dan F 6 .Ini diperlukan untuk mengikat F 1 ke membran .Pengobatan dengan amonia melepaskan OSCP. Sedangkan pengobatan dengan silicotungusate menghilangkan F 6 .
Bagian basa terletak di dalam membran mitokondria bagian dalam. Ini berisi mekanisme translokasi porsi. Ini sesuai dengan FO pada gambar.
Biokimia Mitokondria:
Lindberg dan Ernster (1954) telah memberikan data komposisi kimia mitokondria sebagai berikut: protein 70 sampai 75%, Lipid 25-30% dan RNA 5% dari berat kering.
Tetapi analisis biokimia baru-baru ini menunjukkan komponen-komponen berikut:
(i) Protein
Protein merupakan penyusun utama yang tidak larut dalam air. Membran pembatas luar mitokondria mengandung kurang dari 10 persen dari total protein. Ada sekitar 14 protein berbeda yang memiliki berat molekul dari 12.000 hingga 22.000.
Membran bagian dalam mengandung sekitar 60% protein yang memiliki berat molekul bervariasi dari 10.000 hingga 90.000. Komposisi protein membran mitokondria tidak sepenuhnya diketahui.
Lokalisasi Enzim, Diperoleh dalam Studi Fraksinasi
|
|
Fraksi mitokondria |
Enzim terletak |
|
1. |
Membran luar. |
Monoamine oksidase, “Rotenone-insensitive “NADH-cyto-chrome-C-reductase, Kynurenine hydro xylase, Fatty acid CoA ligase, Glycerophosphate acyl transferase, Nucleoside diphosphokinase, |
|
2. |
Ruang antar membran. |
Adenylate kinase, Nucleoside diphos phokinase, Nucleoside monophosphokinase, |
|
3. |
Membran dalam. |
Enzim rantai pernapasan, β-Hydroxybutyrate dehydrogenase, Ferrochelatase, Carnitine palmityl-transferase, Sistem oksidasi asam lemak, Xylitol dehydrogenase, |
|
4. |
Matriks |
Malate, isocitrate dan Glutamat dehydrogenases, Fumarse, Aconitase, Sitrat sintetase. Ornithine-Carbony 1 transferase, Sistem oksidasi asam lemak, Piruvat karboksilase, |
(ii) Lipid :
Lipid membentuk sekitar 1/5 dari berat membran. Itu hadir hampir seluruhnya dalam bentuk molekul yang dikenal sebagai fosfolipid. Telah dilaporkan oleh Meluick dan Packer pada tahun 1971 bahwa fraksi membran luar memiliki kandungan lipid 40% dibandingkan dengan 20% pada membran dalam.
(iii) Enzim :
Sekitar 70 enzim dan 12 koenzim telah dikenali di mitokondria. Enzim terletak di daerah membran non-air sebagai susunan padat, dengan mungkin sebanyak 5.000 hingga 20.000 kumpulan seperti itu dalam satu mitokondria hati atau jantung.
(iv) DNA mitokondria:
Baru-baru ini DNA juga dilaporkan dari mitokondria. DNA mitokondria beruntai ganda seperti DNA inti. Setiap mitokondria dapat mengandung satu atau lebih molekul DNA tergantung pada ukurannya, jika mitokondria lebih besar dari itu mungkin memiliki lebih banyak molekul DNA. DNA memiliki bentuk lingkaran.
DNA mitokondria berbeda dari DNA inti dalam beberapa hal. Kandungan guanin dan sitosin lebih tinggi pada DNA mitokondria, dan akibatnya kepadatan apung juga lebih tinggi. Jumlah informasi genetik yang dibawa oleh DNA mitokondria tidak cukup untuk memberikan spesifikasi untuk semua protein dan enzim yang terdapat dalam organoid ini. Kemungkinan yang paling mungkin adalah bahwa kode DNA mitokondria untuk beberapa protein struktural.
Mitokondria ragi telah terbukti mengandung DNA polimerase (Winters Berger, 1966) dan baru-baru ini Kalfl968 telah berhasil mengisolasi enzim dari mitokondria hati tikus; DNA polimerase mitokondria tampaknya terlibat dalam replikasi DNA daripada perbaikan (Karol dan Simpson 1968) dan memiliki sifat yang berbeda dari enzim inti.
Ini termasuk persyaratan yang berbeda untuk ion logam (Meyr dan Simpson 1968). DNA polimerase mitokondria ragi tampaknya lebih kecil dari padanan nuklirnya, dan aktif pada berbagai tahap siklus sel (Iwashima dan Rabinowitz, 1969). Bukti visual yang menunjukkan apa yang tampak sebagai DNA mitokondria hati tikus dalam proses replikasi telah disajikan oleh Kirschner, Wolsten Holme dan Gross (1968).
DNA mitokondria tampaknya tidak memiliki histon yang terkait dengannya seperti halnya DNA inti dari organisme yang lebih tinggi. Dalam hal ini DNA mitokondria mirip dengan DNA bakteri.
(v) RNA mitokondria (mt RNA):
South dan Mehlear (1968) menyatakan bahwa jumlah mt-RNA sekitar 10 sampai 20 kali lipat dari mt-DNA. Semua jenis RNA telah diidentifikasi dalam mitokondria. Bukti saat ini menunjukkan secara meyakinkan bahwa mitokondria mengandung set lengkap nRNA (Wintersberger dan Tuppy. 1965), sintesis RNA aminoasil (Barnett, Braun dan Epler, 1967), serta RNA ribosom (Rogers, Preston, Titchener dan Linnane, 1967) .
Semua komponen ini berbeda dari komponennya masing-masing di groundplasma. Keberadaan m-RNA, yang ditranskripsi dari DNA mitokondria masih belum pasti. Namun, ada otoritas yang menyarankan kehadirannya. RNA ribosom dikodekan oleh DNA mitokondria dan dengan demikian tampaknya disintesis di dalam mitokondria oleh sistem RNA polimerase yang bergantung pada DNA mitokondria (Wintersberger, 1964).
(vi) Ribosom mitokondria:
Mitokondria tampaknya mengandung ribosom yang diameternya lebih kecil daripada ribosom sitoplasma {Swift, 1965) dan mitokondria Ragi mengandung spesies RNA 23 detik dan 16 detik (Winter Berger 1966) yang akan sesuai dengan ribosom 70 detik dari jenis bakteri daripada SOS ribosom sitoplasma.
Partikel mirip ribosom dengan nilai sedimentasi 8IS dan 55S juga telah dilaporkan, dan tingkat degradasi yang dialami partikel selama isolasi belum jelas. Agregasi ribosom seperti polisom telah diamati di bagian mitokondria ragi oleh Vignais, Huet dan Andre pada tahun 1969.
Berat molekul tinggi dan spesies RNA yang terkait dengan mitokondria yang berbeda dalam nilai sedimentasi dari RNA ribosom sitoplasma telah dilaporkan dalam sel Ragi, Neurospora dan He-La. Ribosom mitokondria membutuhkan konsentrasi ion Mg ++ yang lebih tinggi untuk menjaga integritasnya daripada ribosom sitoplasma.
Sintesis protein:
Secara umum, mitokondria dapat mengkode dan mensintesis protein, tetapi DNA yang ada di dalamnya tidak cukup untuk mengkode semua protein. Disarankan bahwa mitokondria dapat mensintesis protein yang bersifat struktural (sitokrom oksidase), tetapi banyak protein jika tidak semua, dari protein terlarut dari matriks serta protein dari membran luar dan sejumlah protein yang terletak di criptae (Borst, 1972) berada di bawah kendali nuklir.
DNA dari protein yang dikodekan oleh DNA inti, secara umum disepakati bahwa m-RNA yang berasal dari nukleus diterjemahkan dalam sitoplasma, dan protein yang dihasilkan kemudian diangkut ke dalam mitokondria. Bagaimana protein ini memasuki mitokondria?
Dua metode telah diusulkan:
(1) Prekursor memasuki mitokondria dan di dalamnya diubah menjadi produk akhir, sehingga mempengaruhi aliran material searah ke dalam mitokondria.
(2) Ada sintesis vesikel lipoprotein yang bergabung dan bergabung dengan mitokondria yang sedang tumbuh.
Fungsi:
1. Peran Mitokondria dalam Pembentukan Kuning Telur:
Ada banyak penyelidikan, yang laporannya mengungkapkan bahwa, mitokondria membantu pembentukan kuning telur dalam sel telur yang sedang berkembang. Kajian pertama di bidang ini dilakukan oleh Loyez (1911) dan terakhir mungkin oleh MDL Srivastava (1965), dengan bantuan mikroskop cahaya. Bukti yang dikemukakan tergantung pada topografi, dan hubungan ukuran, dan reaksi pewarnaan mitokondria dan kuning telur protein awal.
Dalam sitologi modern dengan penyelidikan mikroskop elektron, era baru telah dimulai dan studi tentang pembentukan kuning telur tidak jauh dari mikroskop elektron. Dengan bantuan mikroskop elektron Farvard dan Carasso (1958) sampai pada kesimpulan bahwa mitokondria berubah menjadi butiran kuning telur di dalam telur Planorbis coneus.
Perubahan struktural utama yang mereka amati dalam mitokondria adalah sebagai berikut:
(i) Krista menjadi tidak teratur di beberapa membran, tetap konsentris ke membran luar sebelum jatuh sepenuhnya.
(ii) Dalam matriks muncul butiran-butiran kecil yang tersebar lebih dulu, tetapi akhirnya terkumpul menjadi massa dalam pola yang teratur.
2. Selama Pembelahan Sel dan Spermatogenesis:
Ahli sitologi awal, Benda, Dulberg dan Meves berpendapat bahwa mitokondria juga membelah secara merata selama pembelahan sitoplasma dan mungkin berperan dalam pewarisan. Wilson (1928) berkomentar bahwa tidak sedikit pun bukti yang dihasilkan dari perpaduan antara kondriosom paternal dan maternal. Frederic (1958) secara singkat merangkum berbagai perubahan dalam mitokondria selama pembelahan sel.
Fase pertama menunjukkan penurunan volume total bahan mitokondria; secara bertahap menghentikan gerakannya, penipisan yang jelas, fragmentasi menjadi bola-bola kecil, kehilangan kerapatan optik dan akhirnya asimilasi ke dalam sitoplasma.
Pada fase kedua, ketika sel membelah menjadi dua, mitokondria yang termodifikasi dipisahkan secara pasif, menjadi sel anak: Pada fase ketiga, mitokondria yang termodifikasi disusun kembali dengan penambahan unsur-unsur yang diasimilasi dalam sitoplasma.
Wilson menemukan bahwa di Opisthacanthus, selama spermatogenesis, jumlah mitokondria secara bertahap berkurang. Pollister (1930) menjelaskan dalam Gems bahwa mitokondria menyusun diri menjadi cincin yang terdefinisi dengan baik, tetapi tanpa fusi. Studi mikroskopis modern memberikan kesimpulan tegas mengenai pembelahan mitokondria selama mitosis. Pune (1952) mencatat di korteks adrenal rendah mitokondria sering muncul berpasangan.
Ini menunjukkan bahwa perpecahan, bukannya fusi, sedang terjadi. Dalam transformasi spermatid menjadi spermatozoa banyak perubahan mitokondria yang diamati. Franzen (1956) mengamati pada sperma tersebut, yang ditumpahkan langsung dalam air bahwa mitokondria hadir dalam bentuk umumnya empat atau lima bola di bawah kepala sperma, dan dalam kasus sperma yang dikeluarkan dalam media kental, bola ini berubah menjadi dua pita memanjang seperti mitokondria berfilamen.
Kadang-kadang ini berkembang menjadi ‘bola nebenkern’ yang dapat memanjang dan berputar di sekitar filamen aksial untuk membentuk selubung mitokondria. Yasuzumi (1958) menemukan elektron yang tidak dapat dibedakan dari tetesan lipid.’
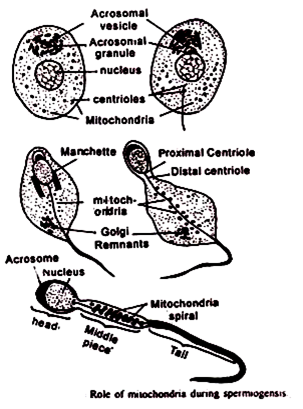
3. Peran mitokondria dalam produksi energi:
Mitokondria memainkan peran yang sangat penting dalam respirasi sel atau produksi energi. Energi diproduksi di dalam sel, sebagian di luar mitokondria dan sebagian besar di dalam mitokondria. Molekul ATP yang diproduksi di sitoplasma non-mitokondria dihasilkan oleh proses yang disebut respirasi anaerob.
4. Peran mitokondria dalam sintesis hem:
Dalam sel hati tikus dan sel darah merah unggas δ-amino levulinate disintesis dari suksin 1 Co-A dan glisin dengan aksi enzimatik δ-amino levulinate synthatase. Enzim ini hadir dalam fraksi mitokondria. δ-amino levulinate adalah perantara penting dalam sintesis porfiri. Jadi mitokondria membantu dalam sintesis hem.
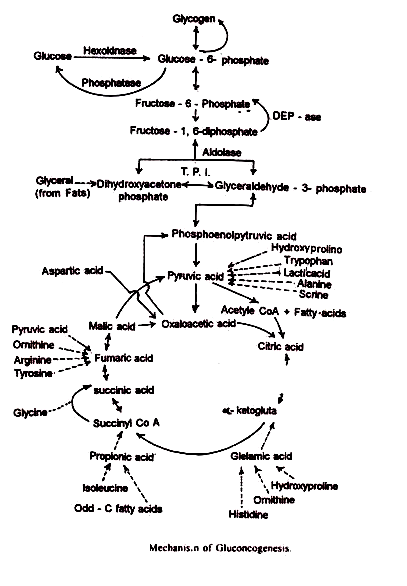
5. Peran dalam glukoneogenesis:
Glukoneogenesis adalah konversi non-karbohidrat menjadi glukosa dari asam piruvat. Diketahui bahwa asam piruvat diubah menjadi asam oksaloasetat dengan adanya karboksilase asam piruvat. Perantara ini dapat keluar dari mitokondria dan diubah menjadi asam fosfoenol piruvat oleh fosfoenol piruvat korboksikinase: Asam fosfoenol piruvat menempati tempat di jalur Embden-Mayerhoff atau jalur glikolitik dari mana jalur tersebut dapat dibalik hingga menjadi glukosa.
Asam amino glukogenik, asam laktat, gliserol dan dalam beberapa kasus propionat dapat, setelah modifikasi yang sesuai, diumpankan pada satu atau titik lainnya dalam siklus Krebs. Asam oksaloasetat dan asam malat dapat keluar dari mitokondria untuk akhirnya diubah menjadi glukosa.
6. Peran mitokondria dalam metabolisme asam amino:
Enzim untuk dominasi oksidatif asam amino terdapat di mitokondria. Ini adalah glutamat dehidrogenase, prolin dehidrogenase, 8-amino levulinat sintetase dll.
7. Peran mitokondria dalam metabolisme lipid:
Mereka mampu mengoksidasi asam lemak. Oksidasi asam lemak membutuhkan oksidasi lengkap asetil Co A dalam siklus Krebs sehingga Co A bebas dapat dihasilkan. Pembalikan oksidasi asam lemak menyebabkan sintesis asam lemak. Selama kelaparan, mitokondria memanfaatkan lemak untuk menghasilkan energi.
Asal Mitokondria:
Pemahaman kita tentang proses dimana mitokondria diproduksi, masih sangat tidak lengkap.
Lehninger (1964) mengklasifikasikan berbagai teori kemungkinan rute asal mitokondria menjadi tiga kelompok utama:
- Pembentukan dari struktur membran lain di dalam sel.
- Pertumbuhan dan pembagian mitokondria yang sudah ada sebelumnya.
- Sintesis de novo dari prekursor submikroskopis.
1. Pembentukan dari struktur membran lain di dalam sel:
Pembentukan mitokondria dengan “mencubit” atau bertunas dari struktur sel yang sudah ada sebelumnya telah disarankan untuk berbagai membran sel termasuk plasmalmma (Robertson, 1959), retikulum endoplasma, amplop nuklir dan kompleks Golgi (Novikoff, 1961) . Tapi dukungan untuk bukti tersebut, dengan tidak adanya data biokimia pendukung, tidak dapat sepenuhnya konklusif.
Bagian dari masalah tidak diragukan lagi terletak pada pengetahuan kita yang terpisah-pisah tentang struktur dan komposisi, dan perbedaan antara membran sel secara umum. Memang kesamaan yang dibahas dalam literatur antara membran mitokondria dan retikulum endoplasma, dapat memberi bobot pada gagasan bahwa mungkin mitokondria terbentuk ketika sitoplasma mendorong ke dalam rongga yang dikelilingi oleh membran internal, yang mereka jepit dan pisahkan dari sistem kontinyu. .
2. Pertumbuhan dan pembagian mitokondria yang sudah ada sebelumnya:
Bukti mikroskopis elektron untuk pembelahan mitokondria dengan fisi, meskipun berlimpah, sulit untuk menilai bahaya memproduksi artefak sangat nyata karena bahan kimia dan fisik yang keras dibawa ke bahan uji selama pemrosesan.
Interpretasi tidak menjadi lebih mudah dengan kemampuan mitokondria untuk mengalami perubahan ekstrim dalam bentuk in vivo yang mungkin terkait atau tidak terkait dengan pembelahan mitokondria. Ada banyak laporan tentang mitokondria yang terhubung satu sama lain melalui jembatan membran yang sempit terutama pada jaringan yang mengalami melopolisasi dengan cepat, dan diperkirakan bahwa angka tersebut dapat mewakili mitokondria pada tahap awal pembelahan.
Dengan mengamati bagian serial dari hati tikus Stempak (1967) mampu menunjukkan bahwa mitokondria “dumb berbentuk lonceng†dapat menjadi bagian dari tubuh berbentuk cangkir. Tubuh seperti itu juga telah diamati pada jaringan pakis yang tumbuh dengan cepat dan mungkin merupakan tahap awal pembelahan.
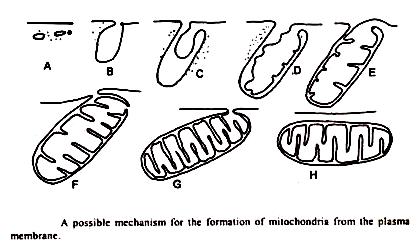
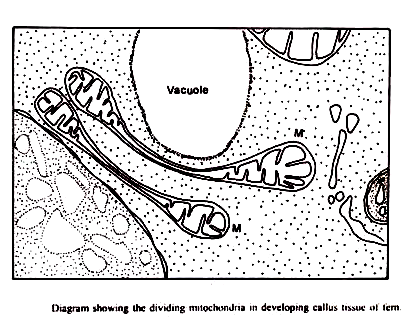
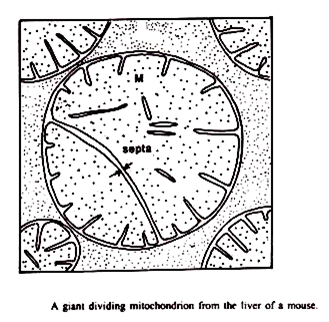
Tahap awal dalam pembelahan mitokondria mungkin melibatkan pemisahan isi mitokondria menjadi dua atau lebih kompartemen. Kehadiran mitokondria dengan “partisi” internal telah terbentuk di beberapa tipe sel (Tandler et. al., 1969) meskipun kemungkinan bahwa mereka adalah manifestasi dari fusi mitokondria tidak dapat dengan mudah dikesampingkan.
Lafontaine dan Allard (1964) telah mempresentasikan pemilihan mikrograf dari mitokondria hati tikus yang menunjukkan apa yang tampak sebagai partisi yang membagi kompleks membran bagian dalam menjadi dua massa, keseluruhannya dikelilingi oleh membran luar yang kontinu Tandler et.al (1969) telah menunjukkan partisi dari mitokondria di hati yang pulih dari defisiensi riboflavin.
3. Lakukan sintesis baru:
Kemungkinan sintesis mitokondria de-novo muncul dengan percobaan di awal abad ini, ketika mitokondria yang mengandung larva terlihat berkembang dari sitoplasma telur bulu babi yang tampaknya telah dibebaskan dari mitokondria dengan sentrifugasi (Novikoff, 1961).
Dengan menggunakan kekuatan pemecahan yang lebih besar dari mikroskop elektron, kemudian ditunjukkan, bahwa mitokondria tidak dapat dikeluarkan dengan sentrifugasi telur (Lansing, Hiller, Rosenthall 1952). Dalam percobaan, mitokondria mungkin telah hadir di “ujung sentripetal” sel telur, dan mitokondria ini dapat berfungsi sebagai prekursor dalam produksi mitokondria selanjutnya.
Pada uraian di atas telah diuraikan beberapa pandangan mengenai pembentukan mitokondria dalam sitoplasma berbagai jenis sel. Tetapi mungkin tidak bijaksana untuk mengelompokkan bukti yang sesuai dengan satu atau beberapa metode terbatas yang dapat digunakan mitokondria untuk bereplikasi.
Situasi sebenarnya mungkin rumit, dan mungkin saja metode replikasi yang berbeda terjadi di jaringan yang berbeda, dan pada tahap perkembangan yang berbeda. Seseorang dapat membayangkan mitokondria awal terbentuk dari struktur membran dalam konsentrasi embrio mitokondria yang berkembang di sekitar membran nukleus telah dicatat dalam jaringan embrionik dari beberapa filum (North dan Poliak, 1961) dan pembentukan mitokondria dari membran ini dapat melibatkan transfer informasi genetik nuklir penting untuk pertumbuhan dan perkalian mitokondria berikutnya dengan pembagian.
Penggandaan mitokondria kemudian dapat dilanjutkan dengan penggabungan molekul besar yang telah dibuat sebelumnya dan asosiasi molekul, dengan pembagian oleh fisi, ketika mitokondria mencapai tahap kritis.
4. Asal Mitokondria Prokariotik:
Fakta bahwa mitokondria dapat tumbuh, membelah, dan mampu bermutasi mendukung pandangan lama bahwa mitokondria berasal dari inangnya. Bakteri akan berasal mitokondria dan ganggang hijau biru, kloroplas.
Ada banyak homologi antara mitokondria dan bakteri. Pada bakteri, sistem transpor elektron terlokalisasi di membran plasma yang dapat dibandingkan dengan membran dalam mitokondria.
Beberapa bakteri bahkan memiliki proyeksi membran yang memanjang dari membran plasma (Fitz-James, 1960) yang sebanding dengan krista mitokondria karena keduanya mengandung rantai pernapasan (Salton dan Chapman, 1962).
Membran bagian dalam dan matriks, telah didalilkan dapat mewakili simbion asli yang dapat tertutup di dalam membran asal seluler (ER). Selanjutnya DNA mitokondria berbentuk sirkular, bereplikasi dan membelah seperti bakteri.
Ribosom juga ditemukan yang, bagaimanapun, lebih kecil dari bakteri. Pada mitokondria dan bakteri, sintesis protein dihambat oleh kloramfenikol. Dari kesamaan ini, seseorang dapat dengan mudah memahami mitokondria sebagai berevolusi dari prokariota kuno (Swift, 1965), yang memiliki semua atribut organisme aerobik yang independen.
Namun, dengan adaptasi dalam waktu yang lama, ia menjadi simbion yang esensial dan bergantung, kehilangan sebagian identitasnya ke sel, dan sebaliknya, sel inang kehilangan sebagian fungsinya, sekarang diturunkan dari endosimbion atau mitokondria. Akibatnya, keduanya menjadi simbion wajib satu sama lain.
Hipotesis simbion tentang asal usul mitokondria dan plastida ini telah mencapai popularitas yang luas, tetapi tidak semua ahli biologi menerimanya. Raff dan Mahler (1972) menyimpulkan bahwa, sementara teori simbiosis mungkin menyenangkan secara estetika, namun tidak menarik.
Mereka menyajikan banyak bukti dan mengusulkan bahwa mitokondria muncul dengan blebbing ke dalam dari membran plasma, entah bagaimana dengan akuisisi membran luar, dan dengan akuisisi tambahan genofor DNA dari DNA proto-eukariota di mana evolusi mitokondria muncul. Borst (1972) mengusulkan teori episode dan mengira bahwa DNA mitokondria meninggalkan DNA ‘nuklir’ dengan semacam amplifikasi untuk dipetakan dalam membran yang berisi rantai pernapasan.



