Proses Metabolisme Sekunder Pada Tumbuhan!
Tumbuhan tidak memiliki mekanisme untuk pembuangan limbah metabolisme mereka. Produk limbah ini terakumulasi dalam sel mereka dan sering menjadi titik awal untuk banyak proses transformasi.
Jalur seperti itu, menghasilkan zat yang tidak penting untuk pemeliharaan jalur metabolisme utama, membentuk apa yang kemudian disebut metabolisme sekunder.
Metabolisme sekunder mengarah pada pembentukan produk sekunder. Jalur metabolisme sekunder diatur secara genetik dan enzimatik. Produk sekunder atau metabolit sekunder meliputi beberapa polisakarida, banyak antibiotik, steroid, karotenoid, alkaloid flavonoid, dll.
Tiga jenis produk sekunder dibedakan sehubungan dengan stabilitas metabolisme:
(1) produk akhir yang benar-benar inert secara metabolik,
(2) produk yang stabilitasnya berkaitan dengan keadaan fisiologis atau perkembangan dan
(3) mereka yang memiliki omset terus menerus. Selama metabolisme produk ini, beberapa produk polimer terbentuk: misalnya karet, sporopollenin, lignin.
Pembentukan zat-zat ini dikatalisis oleh fenoloksidase dan peroksidase dan cukup sering, dinding sel merupakan tempat yang disukai untuk pembentukan makromolekul ini. Degradasi produk sekunder menjadi substrat metabolisme primer dipengaruhi oleh sekelompok enzim yang memiliki spesifisitas substrat rendah seperti monoksigenase metilasi dan glukosidase.
Jumlah produk sekunder bervariasi dengan keadaan perkembangan tanaman, musim, iklim, dan bahkan waktu dalam sehari. Ini karena metabolisme sekunder tunduk pada pengaruh beberapa faktor pengatur.
Pentingnya pembentukan produk sekunder tidak terletak pada fakta bahwa mereka berfungsi sebagai bahan sumber untuk regenerasi senyawa yang diperlukan untuk metabolisme primer. Kalaupun itu terjadi, kontribusinya sangat kecil sehingga tidak berdampak pada metabolisme secara keseluruhan.
Tetapi pembentukan produk sekunder memungkinkan berlanjutnya peristiwa metabolisme primer, melalui penghilangan produk dari jalur sintetik yang bersangkutan dari tempat reaksi dan ini jauh lebih penting.
Produk sekunder: Signifikansi ekologis:
Produk sekunder sangat penting dalam pemeliharaan keseimbangan ekologis. Pengaruh timbal balik antara anggota penyusun suatu ekosistem sebagian besar dapat dilacak ke produk sekunder yang dihasilkan oleh spesies komponen sistem.
Hubungan alelopati antara satu tumbuhan dan tumbuhan lainnya sampai batas tertentu ditentukan oleh sifat produk sekundernya. Misalnya, monoterpen yang dilepaskan dari daun Salvia teucophylla dan Artemisia californica akan membuat tanah yang menyerap zat tersebut tidak cocok untuk menghasilkan benih tanaman tahunan.
Produk sekunder: Hubungan tumbuhan – hewan:
Produk tanaman sekunder memiliki pengaruh yang menentukan pada hubungan tanaman-hewan. Mereka melayani tujuan predasi, penyerbukan dan penyebaran benih. Aroma memainkan peran penting dalam menarik serangga yang bersangkutan dengan penyerbukan.
Zat yang terlibat termasuk monoterpen seperti limonene jeruk dan geraniol mawar; amina seperti monoamina dari Arum dan turunan asam benzoat seperti vanilin dari vanila. Warna bunga adalah ekspresi pigmen, yang juga merupakan produk sekunder.
Misalnya, warna putih bunga tertentu karena adanya flavon seperti luteolin; bunga kuning disebabkan oleh karotenoid, betaxanthin dan flavonol kuning dan warna merah dan ungu disebabkan oleh betacyanin dan anthocyanin. Saat matang, buah memiliki lapisan berwarna cerah dan mengeluarkan bau yang khas.
Keduanya memainkan peran penting dalam menarik hewan dan burung dan akhirnya menyebarkan buah dan biji. Warna dan bau berasal dari produk sekunder. Banyak produk tanaman memiliki rasa dan kelezatan yang ditentukan oleh keberadaan produk sekunder. Ini berperan dalam menarik atau menolak herbivora.
Produk sekunder: Resistensi penyakit dan kejadian:
Produk sekunder merupakan bagian dari sistem pertahanan umum tanaman tingkat tinggi terhadap infeksi mikroba. Sebaliknya, parasit membentuk produk sekunder yang membantu mengatasi ketahanan tanaman inang.
Produk sekunder dalam hubungannya dengan Manusia:
Rasa, aroma, warna dan palatabilitas makanan dan minuman disebabkan oleh sifat kimia dari produk sekunder. Banyak obat yang digunakan manusia juga merupakan produk metabolisme sekunder pada tanaman.
Misalnya, D. Sorbital adalah gula alkohol yang digunakan oleh penderita diabetes; reserpin yang merupakan obat antihipersensitif, strychnine yang berfungsi sebagai stimulan sentral dan kina yang merupakan obat antimalaria semuanya merupakan turunan Indole. Narkotika seperti ganja dan ganja adalah monoterpen dari Cannabis sativa.
Minuman kopi, teh, kakao mengandung kafein, teobromin dan teofilin, semuanya purin. Nikotin dari tembakau dan arekaidin dari Areca catechu adalah turunan asam nikotinat. Nilai obat tanaman Solanaceous seperti Datura dan Atropa disebabkan oleh adanya skopolamin dan alkaloid lain di dalamnya.
Berbagai macam produk sekunder berfungsi sebagai bahan baku industri. Minyak biji rami yang digunakan dalam industri cat, minyak jarak yang digunakan dalam pembuatan pelumas dan sabun, lilin yang digunakan dalam pembuatan poles dan pernis, suberin, komponen gabus yang digunakan dalam produksi sumbat dan filter bahan isolasi, semuanya adalah turunan asam lemak.
Tanin yang memiliki aplikasi penting dalam industri kulit adalah polifenol. Karet adalah politerpena yang diproduksi oleh Ficus elastica, Hevea brasiliensis, dll. Minyak atsiri yang banyak digunakan dalam wewangian, kosmetik, dan sediaan pelarut adalah terpenoid.
Mekanisme Gerakan Stomata:
Stomata berfungsi sebagai katup yang dioperasikan turgor karena gerakan membuka dan menutupnya diatur oleh perubahan turgor sel penutup. Setiap kali, sel penjaga membengkak karena peningkatan turgor, pori dibuat di antara mereka. Dengan hilangnya turgor pori-pori stomata tertutup. Stomata umumnya terbuka pada siang hari dan menutup pada malam hari dengan beberapa pengecualian.
Faktor penting yang mengatur pembukaan stomata adalah cahaya, pH tinggi atau CO 2 yang berkurang dan ketersediaan air. Faktor yang berlawanan mengatur penutupan stomata, yaitu kegelapan, pH rendah atau CO2 tinggi dan dehidrasi. Ada tiga teori utama tentang mekanisme pergerakan stomata:
1. Hipotesis Fotosintesis Sel Penjaga (Schwendener, 1881):
Sel penjaga mengandung kloroplas. Pada siang hari kloroplas melakukan fotosintesis dan menghasilkan gula. Gula meningkatkan konsentrasi osmotik sel penjaga. Ini menyebabkan penyerapan air dari sel epidermis di dekatnya. Sel pelindung yang besar membengkok ke luar dan menciptakan pori di antaranya. Namun, aktivitas fotosintesis kloroplas sel penjaga tampaknya dapat diabaikan.
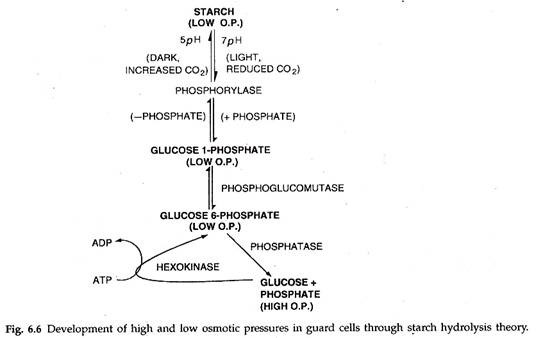
2. Teori Hidrolisis Pati Klasik:
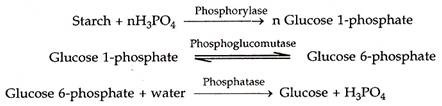
Fitur utama dari teori ini dijabarkan oleh Sayre (1923). Itu telah dimodifikasi oleh Steward (1964). Sel penjaga mengandung pati. Pada konsentrasi karbon dioksida yang rendah (pada pagi hari dicapai melalui fotosintesis oleh mesofil dan sel penjaga), pH sel penjaga meningkat. Ini merangsang enzim fosforilase. Fosforilase mengubah pati menjadi glukosa 1-fosfat. Yang terakhir diubah menjadi glukosa 6-fosfat yang mengalami hidrolisis untuk menghasilkan glukosa dan asam fosfat.
Glukosa meningkatkan konsentrasi osmotik sel penjaga. Karena itu, sel pelindung menyerap air dari sel tetangga, membengkak dan membuat pori di antaranya. Penutupan stomata pada malam hari disebabkan oleh peningkatan kandungan karbon dioksida (karena penghentian fotosintesis) daun. Ini menurunkan pH sel penjaga dan menyebabkan fosforilasi glukosa. Di hadapan fosforilase, glukosa 1-fosfat diubah menjadi pati.
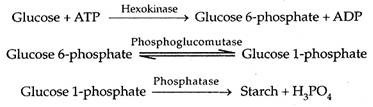
Akibatnya, konsentrasi osmotik sel penjaga turun. Mereka kehilangan air ke sel epidermis yang berdekatan. Dengan hilangnya turgiditas, sel pelindung menyusut dan menutup pori di antaranya.
Keberatan, (i) Glukosa tidak ditemukan pada sel penutup pada saat pembukaan stomata, (ii) Pati <===> Perubahan gula secara kimiawi lambat sedangkan pembukaan dan penutupan stomata cukup cepat. (iii) Perubahan pH sel penjaga yang luas tidak dapat dijelaskan berdasarkan konsentrasi karbon dioksida. (iv) Bawang merah dan beberapa kerabatnya tidak memiliki pati atau polisakarida terkait yang dapat dihidrolisis ke tingkat glukosa, (v) Cahaya biru ditemukan lebih efektif daripada panjang gelombang lain untuk membuka stomata. Hal yang sama tidak dapat dijelaskan dengan teori hidrolisis pati, (vi) Teori hidrolisis pati tidak dapat menjelaskan kenaikan tekanan osmotik yang tinggi yang ditemukan dalam sel penjaga.
3. Hipotesis Pompa Malat atau K + ion (Teori Modern):
Fitur utama dari teori ini dikemukakan oleh Levitt (1974). Menurut teori ini, pH sel penjaga dapat naik karena serapan H + aktif oleh kloroplas sel penjaga atau mitokondria, asimilasi C0 2 oleh mesofil dan sel penjaga. Kenaikan pH menyebabkan hidrolisis pati menjadi asam organik, terutama fosfoenol piruvat. Pati Heksosa Fosfat —> Asam Fosfogliserat —> Fosfoenol Piruvat.
Fosfoenol piruvat juga dapat dibentuk oleh asam piruvat dari jalur pernapasan. Dengan bantuan karboksilase PEP (kasus PEP), ia bergabung dengan CO2 yang tersedia untuk menghasilkan asam oksalat yang diubah menjadi asam malat. Asam malat berdisosiasi menjadi H + dan malat.
H + keluar dari sel penjaga untuk ditukar dengan ion K + . Beberapa ion K + (disertai dengan ion CI- ) juga diserap secara aktif dari sel epidermis yang berdekatan dengan bantuan cAMP, ATP dan sitokinin. Sel penjaga mempertahankan elektronetralitasnya dengan menyeimbangkan K + dengan malat dan CI – .
Dalam keadaan gabungan mereka masuk ke vakuola kecil dan meningkatkan konsentrasi osmotik sel penjaga. Akibatnya, sel pelindung menyerap air dari sel epidermis terdekat melalui endosmosis, membengkak dan membuat pori di antaranya.
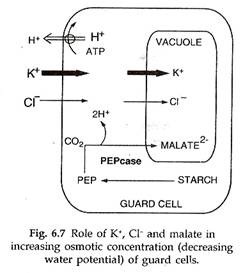
Selama penutupan stomata, ion H + berdifusi keluar dari kloroplas sel penjaga. Ini menurunkan pH sitoplasma sel penjaga. Setiap malat yang ada di sitoplasma bergabung dengan H + untuk membentuk asam malat. Kelebihan asam malat menghambat biosintesisnya sendiri.
CO 2 yang tinggi juga memiliki efek yang serupa. Asam malat yang tidak terdisosiasi meningkatkan kebocoran ion (Jackson dan Taylon, 1970). Akibatnya ion K + terdisosiasi dari malat dan keluar dari sel penutup. Pembentukan asam abcisic (seperti selama kekeringan atau tengah hari) juga mendorong pembalikan H + <===> pompa K + dan meningkatkan ketersediaan H + di dalam sitoplasma sel penjaga.
Hilangnya ion K + menurunkan konsentrasi osmotik sel penjaga dibandingkan dengan sel epidermis yang berdekatan. Hal ini menyebabkan eksosmosis dan karenanya turgiditas sel penjaga berkurang. Ini menutup pori di antara sel penjaga. Secara bersamaan asam organik dimetabolisme untuk menghasilkan pati.



