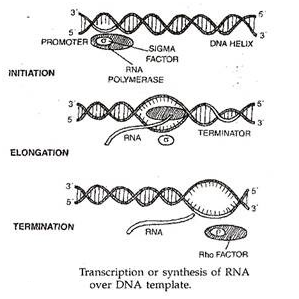
Transkripsi atau Sintesis RNA melalui Templat DNA!
Pada eukariota, transkripsi terjadi pada fase G 1 dan G 2 dari siklus sel di dalam nukleus dan produk transkripsi berpindah ke sitoplasma untuk diterjemahkan. Pada prokariota, transkripsi terjadi dalam kontak dengan sitoplasma karena DNA mereka terletak di sitoplasma.
Transkripsi membutuhkan enzim RNA polimerase yang bergantung pada DNA. Transkripsi segmen DNA memiliki daerah promotor dan terminator. Selain promotor, eukariota juga membutuhkan enhancer.
Faktor terminasi yang disebut faktor Rho(p) yang terdapat dalam DNA diperlukan untuk terminasi transkripsi. Sejumlah faktor lain juga diperlukan untuk pelepasan dupleks DNA, stabilisasi untai DNA yang dilepaskan, pasangan basa, pemisahan dan pemrosesan RNA yang ditranskripsi.
- Aktivasi Ribonukleotida:
Ribonukle otida berbeda dari deoksiribonukleotida karena memiliki gula ribosa dan bukan gula deoksiribosa. Timidin monofosfat digantikan oleh uridin monofosfat. Empat jenis ribonukleotida adalah adenosin monofosfat (AMP), guanosin monofosfat (GMP), uridin monofosfat (UMP) dan cytidine monophosphate (CMP).
Mereka terjadi secara bebas di nukleoplasma. Sebelum transkripsi, nukleotida diaktifkan melalui fosforilasi. Enzim fosforilase diperlukan bersama dengan energi. Ribonukleotida teraktivasi atau terfosforilasi adalah adenosin trifosfat (ATP), guanosin trifosfat (GTP), uridin trifosfat (UTP) dan sitidin trifosfat (CTP).
- Templat DNA:
Pada sinyal tertentu, segmen DNA yang sesuai dengan satu atau lebih sistron menjadi tertekan dan siap untuk ditranskripsi. Setiap segmen transkripsi DNA tersebut memiliki situs inisiasi wilayah promotor, wilayah pengkodean, dan wilayah terminator. Transkripsi dimulai di situs inisiasi dan berakhir di wilayah terminator.
Daerah promotor memiliki situs pengenalan RNA polimerase dan situs pengikatan RNA polimerase. Pembukaan rantai terjadi di wilayah yang ditempati oleh nukleotida TATAAG di sebagian besar prokariota. Enzim yang diperlukan untuk pemisahan rantai adalah unwindases dan protein pengikat untai tunggal. Wilayah terminator memiliki urutan basa poli A atau urutan pallindromik (urutan basa identik yang berjalan berlawanan arah dalam dua rantai DNA).
RNA polimerase (umum pada prokariota dan spesifik pada eukariota) mengikatkan diri ke daerah promotor. Kedua untai DNA mengurai secara progresif dari tempat pengikatan polimerase. Salah satu dari dua untai DNA berfungsi sebagai cetakan untuk transkripsi RNA. Ini disebut master atau untai indra. Transkripsi terjadi dalam arah 5′–> 3′.
- Pasangan Basis:
Trifosfat ribonukleosida yang ada di media sekitarnya datang untuk berbaring berlawanan dengan basa nitrogen dari cetakan DNA (untai indra). Mereka membentuk pasangan komplementer, U berlawanan A, A berlawanan T, C berlawanan G, dan G berlawanan C. Dengan bantuan pyrophos phatase, dua fosfat ekstra hadir pada trifosfat ribonukleosida (ribonukleotida difosfat) terpisah. Energi dilepaskan dalam proses.
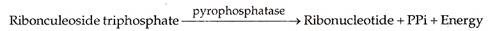
- Formasi Rantai:
Dengan bantuan RNA polimerase, ribonukleotida yang berdekatan yang berada di atas templat DNA bergabung untuk membentuk rantai RNA pada prokariota. Faktor transkripsi sudah berbeda dari RNA polimerase pada eukariota. Saat pembentukan rantai RNA dimulai, faktor sigma (σ) dari RNA polimerase terpisah. RNA polimerase (enzim inti) bergerak di sepanjang cetakan DNA menyebabkan pemanjangan rantai RNA dengan kecepatan sekitar 30 nukleotida per detik. Sintesis RNA berhenti segera setelah polimerase mencapai daerah terminator. Faktor Rho (p) diperlukan untuk ini. Wilayah terminator memiliki sinyal berhenti. Ia juga memiliki 4-8 A-nukleotida.
- Pemisahan RNA:
Pemutusan atau faktor rho memiliki aktivitas ATP-ase (Roberts, 1976). Ini membantu dalam pelepasan rantai RNA yang telah selesai. RNA yang dirilis disebut transkrip primer. Ini diproses untuk membentuk RNA fungsional. Pada banyak prokariota, gen struktural dari fungsi terkait umumnya dikelompokkan bersama dalam operon. Sebuah operon ditranskripsi sebagai satu unit. Unit transkripsi seperti itu adalah mRNA polikistronik. Pada eukariota, unit transkripsi adalah mRNA monokistronik.
- Formasi Dupleks:
Setelah pelepasan transkrip primer, kedua untai DNA membentuk hubungan di antara pasangan basa komplementer. Gyrases, unwindases dan protein SSB dilepaskan. Akibatnya bentuk heliks ganda DNA dilanjutkan.
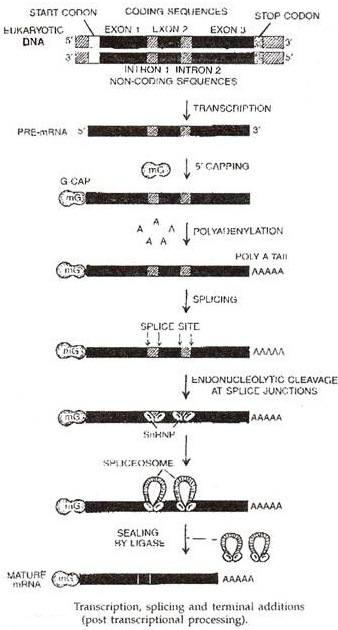
- Pemrosesan Pasca-Transkripsi:
Transkrip primer seringkali lebih besar dari RNA fungsional. Ini disebut hetero gen atau hnRNA terutama dalam kasus mRNA. Pemrosesan pasca transkripsi diperlukan untuk mengubah transkrip primer menjadi RNA fungsional. Ini terdiri dari empat jenis:
(i) Belahan:
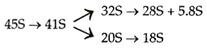
Prekursor RNA yang lebih besar dibelah untuk membentuk RNA yang lebih kecil. Transkrip primer rRNA adalah 45S pada eukariota. Itu dibelah untuk membentuk yang berikut:
Transkrip primer dibelah oleh ribonuclease-P (enzim RNA). Transkrip primer dapat membentuk 5-7 prekursor tRNA.
(ii) Penyambungan:
Transkrip eukariotik memiliki segmen ekstra (intron atau urutan tervening). Urutan pengkodean fungsional disebut ekson. Penyambungan adalah penghilangan intron dan fusi ekson untuk membentuk RNA fungsional. Setiap intron dimulai dengan dinukleotida GU dan diakhiri dengan dinukleotida AG (aturan GU-AG). Mereka dikenali oleh komponen alat penyambung Sn-RNPs (diucapkan sebagai snurps) atau ribonukleoprotein inti kecil (yaitu Ul, U2, U4, U5, dan U6).
Kompleks yang disebut spliceosome terbentuk antara ujung 5′ (GU) dan ujung 3′ (AG) intron. Energi diperoleh dari ATP. Ini menghilangkan intron. Ekson yang berdekatan disatukan. Ujungnya disegel oleh RNA ligase (Gambar 3.13).
Intron bukanlah perkembangan terbaru. Mereka muncul ketika mesin genetik RNA-sentris berada di tempatnya. Oleh karena itu, gen terbelah dan transkripsi terbelah adalah fitur kuno dari sistem genetik. Penyambungan terus menjadi fungsi katalitik yang dimediasi RNA. Banyak lagi proses yang bergantung pada RNA seperti itu yang terungkap.
(iii) Penambahan Terminal (Capping dan Tailing):
Nukleotida tambahan ditambahkan ke ujung RNA untuk fungsi tertentu, misalnya segmen CCA di tRNA, tutup nukleotida di ujung 5′ mRNA atau segmen poli-A (200-300 residu) di ujung 3′ mRNA. Cap dibentuk oleh modifikasi GTP menjadi 7-methyl guanosin atau 7mG.
(iv) Modifikasi Nukleotida:
Mereka yang paling umum di tRNA-metilasi (misalnya, metil sitosin, metil guanosin), deaminasi (misalnya, inosin dari adenin), dihydrouracil, pseudouracil, dll.
Dalam mRNA parkaryotes tidak memerlukan pemrosesan yang rumit untuk menjadi aktif. Selanjutnya, transkripsi dan terjemahan terjadi di wilayah yang sama. Ini menghasilkan awal terjemahan bahkan sebelum mRNA terbentuk sepenuhnya.
Sintesis in vitro RNA pertama kali dilakukan oleh Ochoa (1967).



