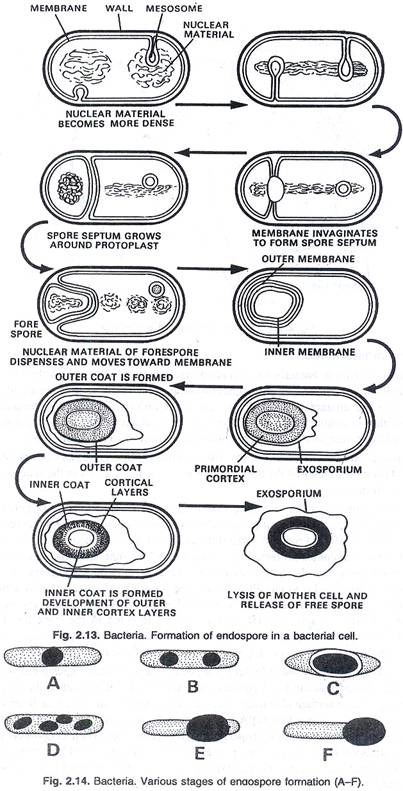
Jenis reproduksi seksual yang terjadi pada bakteri adalah sebagai berikut:
Pengamatan sitologi dan studi genetik menunjukkan sesuatu seperti reproduksi seksual, yang melibatkan peleburan dua sel yang berbeda dan transfer faktor keturunan terjadi pada bakteri walaupun jarang. Rekombinasi genetik terjadi pada bakteri yang telah dipelajari dengan cermat dan mungkin juga terjadi pada spesies lain.

Gambar Courtesy: upload.wikimedia.org/wikipedia/commons/thumb/c/c7/Caduco.jpg/1280px-Caduco.jpg
Salah satu spesies bakteri yang paling intensif dipelajari, Escherichia coli telah terbukti memiliki jenis kelamin bertindak sebagai jantan dan mentransfer informasi genetik melalui kontak langsung dengan betina. Kemampuan untuk mentransfer gen ini diatur oleh faktor kesuburan F+ yang dengan sendirinya dapat ditransfer ke betina, sehingga mengubahnya menjadi jantan.
Sel-sel bakteri vegetatif yang biasa adalah haploid dan dalam reproduksi seksual sebagian atau seluruh kromosom berpindah dari sel jantan ke sel betina, menghasilkan sel, yaitu sebagian atau seluruhnya diploid. Persilangan kemudian terjadi antara kromosom perempuan dan kromosom laki-laki atau fragmen, diikuti dengan proses segregasi yang menghasilkan sel keturunan haploid.
1. Transformasi bakteri:
Transfer genetik pada bakteri juga terjadi melalui transformasi, di mana molekul DNA dari sel donor, ketika dibebaskan oleh disintegrasi, diambil oleh sel penerima lain dan keturunannya mewarisi beberapa karakter sel donor. Ketika strain bakteri yang berbeda ditemukan dalam keadaan campuran baik dalam kultur atau di alam, beberapa keturunan yang dihasilkan memiliki kombinasi karakter dari strain induknya. Fenomena ini dikenal sebagai rekombinasi.
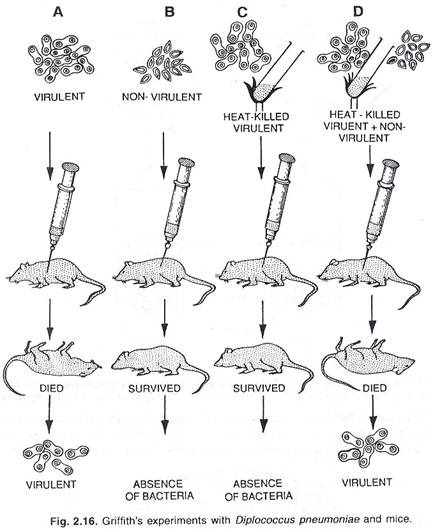
Fenomena transformasi pertama kali dicatat oleh Griffith (1928). Avery, Macleod dan McCarty (1944) mendemonstrasikan bahwa prinsip transformasi menjadi DNA dalam urutan kejadian dalam transformasi bakteri.
Garis penyelidikan yang mengarah pada pemahaman tentang sifat kimia materi genetik muncul dari studi organisme pes Diplococcus pneumoniae. Bakteri ini menyebabkan pneumonia pada pria. Pada tahun 1928, Frederick Griffith menemukan bahwa terdapat dua galur D. pneumoniae, satu membentuk koloni halus yang dilindungi oleh kapsul, dan satu lagi membentuk koloni tidak beraturan atau kasar tanpa kapsul jika ditumbuhkan pada media yang sesuai dalam cawan petri.
Ketika disuntikkan ke tikus (A) hanya sel halus berkapsul (virulen) yang menghasilkan penyakit, tetapi bukan sel kasar non-virulen (B). Di sisi lain ketika panas membunuh sel halus berkapsul (virulen) dicampur dengan sel kasar non-virulen (D) dan kemudian disuntikkan pada tikus, penyakit dihasilkan. Ini menunjukkan bahwa beberapa faktor dari sel halus berkapsul mati, mengubah sel kasar non-virulen hidup menjadi sel berkapsul halus (virulen) hidup, (lihat gbr. 2.16).
Pada tahun 1944, Avery, McCarty dan Macleod mendukung eksperimen Griffith dengan penjelasan molekuler. Mereka menemukan bahwa DNA yang diisolasi dari panas membunuh sel halus, ketika ditambahkan ke sel kasar mengubah karakter permukaannya dari kasar menjadi halus, dan juga membuatnya ganas.
Melalui percobaan ini, ditunjukkan bahwa DNA adalah bahan genetik yang bertanggung jawab untuk menginduksi sifat halus sel dan sifat virulensinya pada tikus. Eksperimen mereka membuktikan bahwa transformasi bakteri melibatkan transfer bagian DNA dari bakteri mati (yaitu, donor) ke bakteri hidup (yaitu, penerima), yang mengekspresikan karakter sel mati, dan dikenal sebagai rekombinan.
Agen penginfeksi virus adalah DNA:
Bakteriofag (virus T 2 ) menginfeksi bakteri Escherichia coli. Setelah infeksi, virus berkembang biak dan fag T2 dilepaskan dengan lisis sel bakteri. Seperti yang kita ketahui, fag T 2 mengandung DNA dan protein. Sekarang muncul pertanyaan, komponen mana yang memiliki informasi untuk memprogram penggandaan lebih banyak partikel virus.
Untuk mengatasi masalah ini Hershey dan Chase (1952) merancang percobaan dengan dua preparat fag T2 yang berbeda . Dalam satu persiapan mereka membuat bagian protein menjadi radioaktif dan dalam persiapan lainnya DNA dibuat radioaktif. Setelah itu biakan E. coli dibuat terinfeksi oleh kedua preparat fag ini. Segera setelah infeksi dan sebelum lisis bakteri, sel-sel E. coli diaduk perlahan di dalam mixer sehingga partikel fag yang menempel terlepas dan kemudian biakan disentrifugasi. Dengan hasil pelet yang lebih berat dari sel bakteri yang terinfeksi menetap di bagian bawah tabung. Partikel virus yang lebih ringan dan partikel yang tidak masuk ke sel bakteri ditemukan di supernatan. Ditemukan bahwa ketika fag T 2 dengan DNA radioaktif digunakan untuk menginfeksi E. coli dalam percobaan, pelet bakteri yang lebih berat juga bersifat radioaktif. Di sisi lain ketika fag T 2 dengan protein radioaktif digunakan, pelet bakteri memiliki radioaktivitas yang sangat sedikit dan sebagian besar radioaktivitas ditemukan di supernatan. Ini
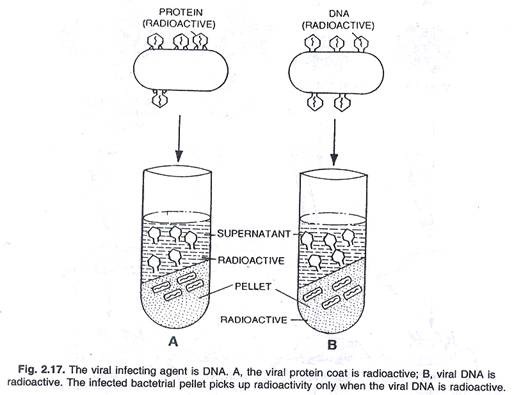
bahwa itu adalah DNA virus dan bukan protein yang mengandung informasi untuk produksi lebih banyak partikel fag T2 sehingga DNA adalah bahan genetik. Namun, pada beberapa virus (misalnya TMV, virus influenza dan virus polio) RNA berfungsi sebagai materi genetik, (lihat gambar 2.17).
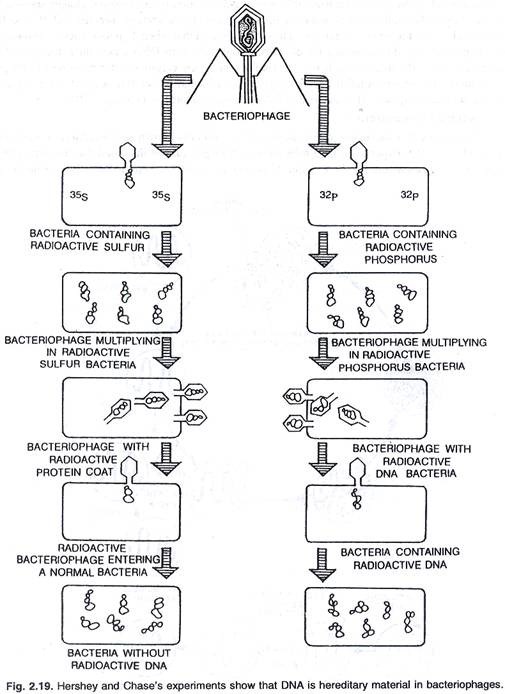
Hershey dan Chase melakukan dua percobaan. Dalam satu percobaan E. coli diberikan dalam media yang mengandung radio-isotop S 35 dan dalam percobaan lain E. coli ditanam dalam media yang mengandung radio-istop P 32 . Dalam percobaan ini sel E. coli dibuat terinfeksi dengan fag T 2 yang dilepaskan dari sel E. coli yang tumbuh dalam media S 35 memiliki S 35 dalam kapsid proteinnya, dan sel dari media P 32 memiliki P 32 dalam DNA-nya.
Ketika fag ini digunakan untuk menginfeksi sel E. coli baru dalam medium normal, sel bakteri yang terinfeksi oleh fag berlabel S 35 menunjukkan radioaktivitas di dinding selnya dan bukan di sitoplasma. Sedangkan bakteri yang terinfeksi fag berlabel P 32 menunjukkan kondisi sebaliknya.
Dengan demikian dapat dikatakan bahwa ketika fag T 2 menginfeksi sel bakteri, kapsid proteinnya tetap berada di luar sel bakteri tetapi DNA-nya masuk ke dalam sitoplasma bakteri. Ketika sel bakteri yang terinfeksi dilisiskan, partikel virus lengkap baru (fag T 2 ) terbentuk. Ini membuktikan bahwa DNA virus membawa informasi untuk sintesis lebih banyak salinan DNA dan kapsid protein. Ini menunjukkan bahwa DNA adalah materi genetik, (lihat gambar 2.19).
2. Transduksi bakteri:
Transfer genetik pada bakteri dicapai dengan proses yang dikenal sebagai transduksi. Eksperimen Lederberg dan Zinder (1952) pada U-tube Salmonella typhimurium menunjukkan bahwa virus bakteri atau fag bertanggung jawab untuk transfer materi genetik dari satu ke yang lain lisogenik dan
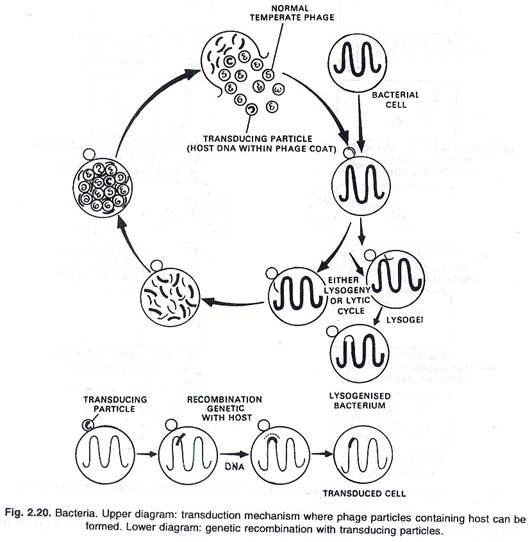
fag litik. Jadi tuan rumah memperoleh genotipe baru. Transduksi telah dibuktikan pada banyak bakteri.
Dalam proses ini, molekul DNA yang membawa karakter herediter dari bakteri donor dipindahkan ke sel penerima melalui agen partikel fag. Dalam proses ini sangat sedikit karakter yang terkait erat yang dapat ditransfer oleh setiap partikel. Jadi bakteriofag membawa perubahan genetik pada bakteri yang bertahan dari serangan fag.
Ketika sel bakteri terinfeksi virus sedang, baik siklus litik atau lisogeni dimulai. Setelah itu, DNA inang terurai menjadi fragmen kecil seiring dengan perbanyakan virus. Beberapa fragmen DNA ini digabungkan dengan partikel virus menjadi satu transduksi. Ketika bakteri melisiskan partikel ini bersama dengan partikel virus normal dilepaskan
ketika campuran transduksi dan partikel virus normal ini dibiarkan menginfeksi populasi sel penerima, sebagian besar bakteri terinfeksi partikel virus normal dan akibatnya lisogeni atau siklus litik terjadi lagi. Beberapa bakteri terinfeksi dengan partikel transduksi, terjadi transduksi dan DNA partikel virus mengalami rekombinasi genetik dengan DNA bakteri. (Lihat gbr. 2.20 dan 2.21).
3. Konjugasi bakteri:
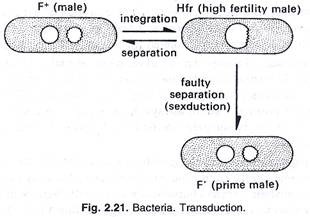
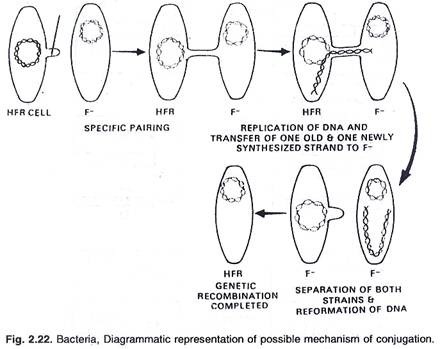
Wollman dan Jacob (1956) telah menjelaskan konjugasi di mana dua bakteri berbaring berdampingan selama setengah jam. Selama periode waktu ini, sebagian materi genetik secara perlahan diwariskan dari satu bakteri yang ditunjuk sebagai jantan ke penerima yang ditunjuk sebagai betina. Ini ditetapkan bahwa materi laki-laki memasuki perempuan dalam rangkaian linier.
Rekombinasi genetik antara sel donor dan penerima terjadi sebagai berikut: DNA Hfr setelah meninggalkan bagian dalam fragmen ke sel penerima kembali membentuk kembali secara melingkar. Pada rekombinasi genetik galur F terjadi antara fragmen donor dan DNA resipien. Transfer gen adalah proses berurutan dan strain Hfr yang diberikan selalu menyumbangkan gen dalam urutan tertentu. DNA donor beruntai tunggal (faktor F) terintegrasi dalam kromosom inang dengan bantuan enzim nuklease, (lihat gambar 2.21 dan 2.22).
Dalam konjugasi bakteri, transfer bahan genetik (DNA) terjadi melalui kontak sel ke sel donor dan sel penerima. Selama proses konjugasi sebagian besar genom ditransfer, sedangkan dalam transformasi dan transduksi hanya sebagian kecil DNA yang ditransfer. Proses konjugasi ditemukan oleh Lederberg dan Tatum (1944) pada Escherichia coli strain tunggal. Konjugasi juga telah didemonstrasikan pada Salmonella, Pseudomonas dan Vibrio.
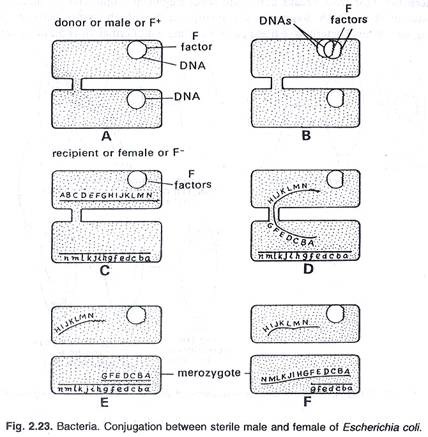
Dalam konjugasi satu arah transfer materi genetik terjadi dari donor ke strain penerima. Strain donor dan penerima selalu ditentukan secara genetik. Strain penerima ditetapkan sebagai F, sedangkan strain donor terdiri dari dua jenis dan ditetapkan sebagai F + dan H fr (rekombinasi frekuensi tinggi). Jika strain menyumbangkan hanya sebagian kecil genomnya disebut F + , dan jika mendonorkan genom dalam jumlah besar disebut H fr. Faktor F + dan H ini disebut episom.
Strain F + dan Hfr ditandai dengan adanya struktur seperti flagel spesifik, yang disebut pilus seks. Pilus seks tidak ada pada strain F + , dan bertanggung jawab untuk perkawinan bakteri. Sex pili F + dan H fr bersentuhan dengan jenis sel kawin yang berlawanan khusus untuk mentransfer materi genetik.
Sex pilus memiliki lubang berdiameter 2,5µm yang cukup besar untuk dilewati molekul DNA secara memanjang. Pada saat memasangkan DNA dari galur H fr (donor) segera ditransfer ke galur F ( penerima). DNA sirkuler sel Hfr terbuka dan bereplikasi tetapi selama transfer, satu untai DNA baru disintesis, sedangkan untai lainnya berasal dari untaian Hfr strain yang sudah ada sebelumnya. Setelah transfer DNA, kedua sel dipisahkan satu sama lain.
H fr DNA setelah memisahkan fragmennya ke sel penerima kembali membentuk kembali secara melingkar. Dalam F – strain rekombinasi genetik terjadi antara fragmen donor dan DNA penerima. Transfer gen adalah proses berurutan, galur H fr yang diberikan selalu menyumbangkan gen dalam urutan tertentu. Jika galur F – dan H fr dibiarkan bercampur dalam suspensi, gen yang berbeda dalam urutan waktu dipindahkan dari genom H fr ke galur F. Gen yang masuk lebih awal, selalu muncul dalam persentase rekombinasi yang lebih besar daripada gen yang masuk belakangan, (lihat gambar 2.22, 2.23 dan 2.24).
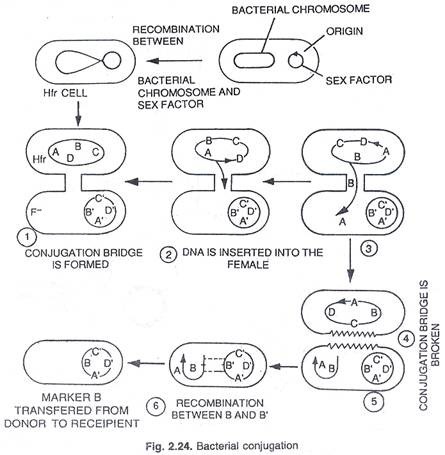
Konjugasi menghasilkan sejumlah rekombinan dalam suspensi sel F + dan H fr. Rekombinan ini bervariasi dalam konstitusi genotipiknya dan juga dalam ekspresi fenotipiknya. Rekombinan ini sama sekali baru dan berbeda dari induknya.



