Baca artikel ini untuk mempelajari tentang sistem limfatik tubuh manusia!
Ini terdiri dari sistem pembuluh tertutup yang bercabang di ruang jaringan di dalam dan sekitar kapiler darah, dan membawa cairan jaringan ke dalam sistem pembuluh darah dengan bertindak sebagai rute alternatif.
Sumber gambar: topknowhow.com/wp-content/uploads/2013/04/lymphatic-system.jpg
Oleh karena itu, sistem limfatik merupakan pelengkap sistem vena. Dalam perjalanannya, limfatik dicegat oleh rantai kelenjar getah bening yang menyaring getah bening dan menambahkan limfosit ke dalam getah bening yang bersirkulasi.
Getah bening:
Cairan jaringan, ketika masuk ke dalam sistem limfatik dikenal sebagai getah bening. Getah bening membawa makromolekul protein dan partikel besar (debu, karbon, bakteri, sel kanker, dll.) menjauh dari ruang jaringan. Konsentrasi protein getah bening yang berasal dari sebagian besar jaringan perifer sekitar 2 gm persen, dan ini hampir identik dengan konsentrasi protein cairan jaringan. Pembentukan getah bening berbanding lurus dengan pembentukan cairan jaringan.
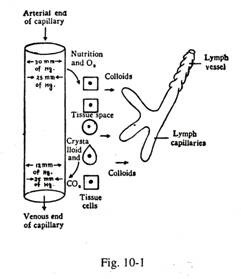
Mekanisme Pembentukan Cairan Jaringan (Gbr. 10-1):
(1) Di ujung arteri kapiler:
Tekanan hidrostatik (gaya penggerak luar) = 30 mm Hg.
Tekanan osmotik protein (daya tarik ke dalam) = 25 mm Hg Tekanan filtrasi (30-25) = 5 mm Hg. Oleh karena itu, sebagian besar kristaloid dan oksigen plasma darah muncul di ruang jaringan untuk nutrisi sel jaringan.
(2) Di ujung vena kapiler:
Tekanan hidrostatik = 12 mm Hg. (pengemudi luar)
Tekanan osmotik protein -25 mm Hg. (menarik ke dalam)
Oleh karena itu, sebagian besar kristaloid dan molekul mikro dari koloid cairan jaringan bersama dengan karbon dioksida diserap kembali melalui dinding en dotelial semipermeabel dari ujung vena kapiler. Tetapi cairan jaringan sementara itu mengandung makromolekul koloid dari metabolit jaringan; kadang-kadang partikel dapat ditambahkan ke cairan.
Bahan-bahan cairan jaringan ini (koloid dan partikel ) diserap ke dalam sistem limfatik melalui dinding endotel kapiler limfatik yang sangat permeabel. Sekitar sepersepuluh dari cairan jaringan, kaya kandungan protein, memasuki kapiler getah bening. Salah satu fungsi penting dari saluran limfatik adalah untuk mempertahankan konsentrasi protein yang rendah dari cairan jaringan, karena ini adalah satu-satunya jalan dimana kelebihan protein dapat kembali ke sistem peredaran darah.
Fungsi Sistem Limfatik:
(1) Membantu sistem vena untuk mengalirkan komponen koloid dan partikel dari cairan jaringan;
(2) Membantu penyerapan lemak hasil cerna langsung ke cisterna chyli. Dua pertiga lemak diserap oleh sistem limfatik;
(3) Menyaring partikel dan agen berbahaya getah bening melalui aksi fagositik sel makrofag kelenjar getah bening;
(4) Menghasilkan limfosit oleh folikel limfatik kelenjar getah bening;
(5) Menghasilkan zat kekebalan (antibodi ) oleh sel plasma kelenjar getah bening;
(6) Dalam kondisi patologis, itu menyediakan saluran untuk penyebaran infeksi atau sel ganas. Serum antioksidan yang disuntikkan ke dalam ruang jaringan diserap oleh limfatik; bisa ular kobra diserap sebagian oleh kapiler darah dan sebagian lagi oleh kapiler limfe.
Komponen Sistem Limfatik:
(A) Pembuluh limfe
(1) Pembuluh getah bening
(2) Pembuluh limfa yang tepat
(3) Saluran getah bening terminal
(B) Jaringan limfoid
(1) Folikel limfatik primer
(2) Kelenjar getah bening
(3) Kelenjar hemolimf
(4) Timus
Pembuluh Limfe
Kapiler Getah Bening (Gbr. 10-1):
Ini dimulai secara membabi buta di ruang jaringan di sekitar kapiler darah dan berkomunikasi secara bebas dengan kapiler getah bening yang berdekatan. Kapiler tidak memiliki katup, dilapisi oleh en dothelium pipih yang tidak memiliki lamina basal dan sel pericyte yang pasti, dan berlabuh ke serat jaringan ikat. Oleh karena itu, kapiler limfe tidak dapat kolaps selama peningkatan tekanan cairan jaringan. Sel-sel endotel kapiler tidak terhubung satu sama lain melalui persimpangan ketat.
Tepi sel-sel en dothelial tumpang tindih sedemikian rupa sehingga katup penutup terbentuk di antara sel-sel yang berdekatan sehingga cairan jaringan dapat masuk ke dalam kapiler getah bening, tetapi aliran balik getah bening kembali ke ruang jaringan dicegah oleh katup penutup. (Gbr. 10-2). Dinding kapiler sangat permeabel terhadap makromolekul koloid. Getah bening yang muncul di kapiler bersifat multifugal. Kapiler getah bening tidak ada di area berikut:
(a) Struktur avaskular—epidermis, kornea , dan kartilago hialin artikular;
(b) Pulpa limpa dan sumsum tulang;
(c) lobulus hati dan unit paru;
(d) Otak dan sumsum tulang belakang.
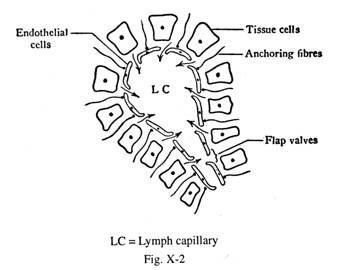
Faktor-faktor yang membantu cairan jaringan masuk ke dalam kapiler getah bening:
(1) Tekanan filtrasi cairan jaringan—Baru-baru ini ditemukan bahwa tekanan cairan jaringan bersifat sub-atmosfer hingga sekitar -6 mm sampai -7 mm Hg., dan tekanan osmotik koloid cairan jaringan diberikan oleh tekanan rendah konsentrasi protein sekitar 5 mm Hg. (Konsultasikan dengan Fisiologi Medis – Guyton) Meskipun ada tekanan negatif, cairan jaringan memasuki kapiler limfatik melalui hisapan yang dibuat oleh pompa limfatik yang mendorong aliran getah bening. Tekanan cairan jaringan negatif seperti itu membantu menahan berbagai jaringan tubuh. Ketika tekanan cairan jaringan menjadi positif, edema berkembang.
(2) Difusi kristaloid melalui membran semipermeabel;
(3) Koloid memasuki lumen kapiler melalui katup flap di antara sel-sel endotel dengan semacam tindakan hisap.
Pembuluh Getah Bening:
Pembuluh getah bening tampak seperti manik-manik, dilengkapi dengan katup, dan lebih banyak dari vena. Kapal terdiri dari set dangkal dan dalam. Pembuluh superfisial terletak di jaringan areolar subkutan dan menyertai vena. Pembuluh dalam terletak di bawah penutup fasia dalam dan menyertai arteri.
Struktur pembuluh getah bening berukuran sedang:
(a) Tiga lapisan dari dalam ke luar – Tu nica intima dilapisi oleh endotelium. Tunika media terdiri dari otot polos yang tersusun melingkar. Tunika eksternal terdiri dari jaringan fibrosa.
(b) Kapal dilengkapi dengan banyak katup yang semilunar dan disusun berpasangan; tepi bebas dari katup diarahkan sepanjang aliran getah bening. Di bagian proksimal dari katup, dindingnya melebar; karenanya manik-manik dalam penampilan.
Keunikan kapal:
(1) Getah bening mengalir dalam satu arah saja (unifugal), dipandu oleh katup.
(2) Aliran mundur dapat terjadi jika pembuluh terhalang, oleh pemisahan katup.
(3) Kadang-kadang pembuluh getah bening berakhir ke kelenjar getah bening yang jauh dan melewati kelenjar getah bening langsung melalui pleksus perikapsular. Setetes getah bening di dalam kapiler dapat mengalami perjalanan yang membingungkan dan bervariasi melalui pleksus rumit dari kapiler getah bening tanpa katup, tergantung pada kondisi setempat. Tapi begitu getah bening muncul di pembuluh yang tepat yang dilengkapi dengan katup, rutenya menjadi tidak dapat diubah.
Kadang-kadang penjelasan drainase limfatik seperti yang dijelaskan oleh berbagai pekerja tidak sepenuhnya sesuai satu sama lain. Mungkin semua pekerja benar dalam uraiannya masing-masing dalam keadaan yang berbeda. Perlu dicatat bahwa drainase limfatik organ tertentu dalam kondisi patologis berbeda dengan drainase saat organ sehat.
Saluran Limfe Terminal:
Ini adalah saluran toraks dan saluran limfatik kanan, dan masing-masing berakhir menjadi vena brachio-cephalic kiri dan kanan. Saluran toraks mengalirkan getah bening dari seluruh tubuh kecuali sisi kanan kepala dan leher, tungkai kanan atas, dinding toraks kanan, paru-paru kanan, sisi kanan jantung dan sebagian permukaan cembung hati. Laju aliran getah bening melalui saluran toraks pada orang yang sedang istirahat adalah sekitar 100 ml per jam.
Faktor yang mengatur aliran getah bening:
saya. Tekanan filtrasi cairan jaringan.
- Tindakan memijat otot rangka ; ini adalah faktor yang sangat penting.
aku aku aku. Pulsasi arteri yang ditransmisikan.
- Katup mengubah bejana menjadi segmen kecil, dan mengarahkan aliran.
v.Gravitasi.
- Tindakan hisap diafragma dan tekanan negatif pada vena brachiocephalic.
Jaringan Limfoid:
Ini adalah jaringan ikat yang dimodifikasi dan terdiri dari struktur dan sel pendukung. Struktur pendukung dibentuk oleh jaringan pleksiform serat retikuler yang mengandung banyak sel di celahnya. Sel terdiri dari dua jenis—tetap dan bebas.
Sel tetap adalah sel retikuler yang melekat pada serat retikuler dan berfungsi fagositik. Sel retikuler dikenal sebagai sel litoral, dan bertindak sebagai sel induk dari mana sel bebas berasal. Sel-sel bebas terdiri dari limfoblas, limfosit, dan sel plasma, dan menempati celah antara serat retikuler.
Sel plasma terbentuk dari limfosit B, ketika sel tersebut diinduksi oleh antigen ke dalam sel yang kompeten secara imunologis yang dikenal sebagai imunoblas. Imunoblas digantikan oleh plasmoblas, proplasmatosit, dan sel plasma. Setiap sel plasma menghasilkan antibodi sirkulasi spesifik untuk antigen tertentu.
Setelah sel plasma menghasilkan satu jenis antibodi, secara permanen berkomitmen untuk menghasilkan hanya jenis antibodi yang spesifik untuk antigen tersebut di atas. Limfosit terdiri dari dua varietas—sel yang bergantung pada timus, limfosit-T, yang berkaitan dengan imunitas yang dimediasi sel dan reaksi hipersensitivitas lambat; sel independen timus, limfosit B, yang menghasilkan antibodi humoral melalui sel plasma. Limfoblas berasal dari limfosit T dan В, dan membesar pada stimulasi antigenik sebelum membelah menjadi limfosit kecil.
Antigen kita, zat asing yang masuk ke dalam tubuh menghasilkan respons imunologis dari inang untuk perlindungan diri dengan menonaktifkan atau menghancurkan zat asing. Responnya mungkin dalam bentuk imunitas seluler yang dimediasi terutama oleh limfosit T (sel T), atau imunitas humoral yang dihasilkan oleh sel plasma yang berasal dari limfosit Ð’ (sel Ð’), atau paling sering dengan kedua metode tersebut. Antigen harus asing dan tubuh harus mengenalinya sebagai benda asing.
Pengenalan antara antigen diri dan non-diri berkembang dalam kehidupan intra-uterus mungkin dengan bantuan kelenjar timus. Antigen dapat terdapat dalam sel utuh (misalnya bakteri, sel tumor) atau dalam makromolekul protein, polisakarida, atau nukleoprotein. Sebagai aturan umum, molekul yang lebih besar dari 10.000 berat molekul cukup imunogenik. Senyawa kimia dengan berat molekul lebih kecil dapat bertindak sebagai antigen ketika menempel pada molekul besar; molekul yang lebih kecil seperti itu dikenal sebagai hapten.
Spesifisitas respon imun dikendalikan oleh unit molekul yang relatif kecil, penentu antigenik dari antigen. Penentu antigenik untuk protein terdiri dari 4-6 asam amino dan untuk polisakarida adalah unit monosakarida. Sel bakteri yang memiliki banyak determinan antigenik akan menimbulkan spektrum respon seluler dan humoral yang luas. Antigen penghasil respon imunologi dikenal sebagai imunogen.
Antigen tersebut berasal dari mikroorganisme (virus, bakteri, jamur, parasit dan cacing), dari sel jaringan yang berbeda secara genetik atau dari beberapa sel tubuh inang yang menjadi bermusuhan dan menghasilkan reaksi autoimun. Sel kanker juga bertindak sebagai antigen.
Antibodi atau imunoglobulin adalah protein plasma makro-molekul yang bersirkulasi yang disekresikan oleh sel plasma inang. Yang terakhir berasal dari limfosit B yang diaktifkan oleh antigen untuk menghasilkan antibodi spesifik terhadap antigen tersebut melalui sel plasma dengan tujuan menonaktifkan atau menetralkan aksi antigen.
Berat molekul antibodi bervariasi antara 150.000 dan 950.000. Setiap molekul antibodi terdiri dari empat rantai polipeptida, sepasang rantai berat (H) identik, dan sepasang rantai ringan (L) identik, yang diikat oleh ikatan disulfida.
Seluruh molekul menyerupai huruf ‘Y’. Masing-masing tungkai yang menyimpang terdiri dari satu rantai berat dan satu rantai ringan, dan bertindak sebagai tempat pengikatan antigen . Batang ‘Y’ dibentuk oleh aposisi dua rantai berat saja; itu bertindak sebagai situs pengikat reseptor dan menyediakan situs untuk lampiran komplemen.
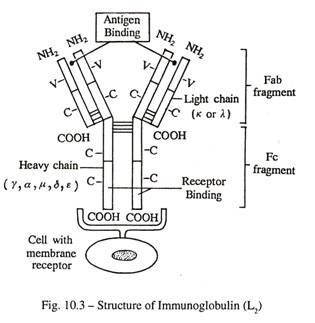
Dua jenis rantai-L, Kappa (к) dan Lambda (λ) telah dibuktikan pada manusia, berdasarkan urutan asam amino daerah konstan. Molekul antibodi tertentu selalu mengandung rantai к atau λ yang identik, tidak pernah campuran keduanya (Gbr. 10-3).
Lima kelas rantai-H telah ditemukan pada manusia, berdasarkan perbedaan struktural di daerah konstan . Berbagai bentuk rantai H, yang ditunjuk γ, α, µ, δ, dan e, ditemukan di IgG, IgA, IgM, IgD dan IgE.
Setiap rantai polipeptida terdiri dari sejumlah loop atau domain dengan ukuran konstan yang dibentuk oleh ikatan disulfida intra-rantai. Domain N-terminal dari setiap rantai menunjukkan lebih banyak variasi dalam urutan asam amino daripada yang lain dan ditetapkan sebagai domain variabel; wilayah lain disebut domain konstan di setiap rantai. Saat diobati dengan enzim papain, molekul imunoglobulin pecah menjadi tiga fragmen dengan ukuran yang sama; dua fragmen Fab (pengikatan antigen) yang mencakup satu rantai ringan utuh dan domain V H dan C H I dari rantai berat; satu fragmen Fc (dapat dikristalisasi) yang terdiri dari bagian С-terminal dari rantai berat. Daerah Fc bereaksi dengan reseptor spesifik dari banyak sel yang berbeda dan dengan fiksasi komplemen .
Kelas Imunoglobulin:
Lima Kelas diakui dalam manusia:
1.Igg:
Ini adalah kelas yang paling melimpah, merupakan sekitar 75% dari total imunoglobulin serum dan ada dalam bentuk monomer. Ini adalah satu-satunya imunoglobulin yang melintasi penghalang plasenta dan melindungi bayi baru lahir dari infeksi.
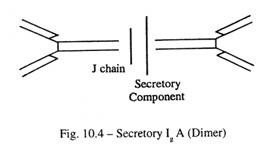
- IgA (Gbr. 10-4):
Ini adalah imunoglobulin utama dalam sistem kekebalan mukosa, dan hadir dalam air liur, air mata, sekresi bronkial, mukosa hidung, cairan prostat, sekresi vagina dan sekresi mukosa usus kecil.
IgA sekretori ditemukan dalam bentuk dimer, dan terdiri dari dua molekul IgA monomer yang disatukan oleh protein J dan digabungkan dengan protein lain, komponen sekretori. Monomer IgA dan protein J disekresikan oleh sel plasma di selaput lendir yang melapisi saluran pencernaan, pernapasan, dan saluran kemih; komponen sekretorik disintesis oleh sel epitel mukosa.
Sekretori IgA resisten terhadap enzim pencernaan proteolitik. IgA biasanya ada dalam serum dalam bentuk monomer dan polimer, yang merupakan sekitar 15% dari total imunoglobulin serum.
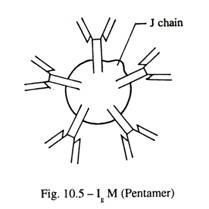
- IgM (Gbr. 10-5):
Ini merupakan 10% dari serum imunoglobulin dan ada sebagai pentamer dengan berat molekul sekitar 900.000. Ini adalah imunoglobulin dominan dalam respon kekebalan awal, dan bersama dengan IgD, ditemukan pada permukaan B-limfosit.
IgM dan IgD menunjukkan bentuk terikat membran dan sirkulasi. IgM dan IgD yang terikat membran berfungsi sebagai reseptor untuk antigen spesifik, yang membantu proliferasi dan diferensiasi limfosit B, menghasilkan sel plasma yang mensekresi antibodi. IgM juga mengaktifkan sistem komplemen, yaitu sekelompok protein plasma yang mampu menghasilkan lisis sel, termasuk bakteri.
4.IgE:
Biasanya ada sebagai monomer dan memiliki afinitas yang besar untuk reseptor yang terletak di membran plasma sel mast dan basofil (antibodi sitofilik). Segera setelah disekresikan oleh sel plasma, IgE menempel pada sel-sel ini dan hampir menghilang dari plasma darah dan hanya merupakan 0,004% dari total imunoglobulin serum.
Ketika antigen spesifik yang menghasilkan antibodi IgE ditemui lagi, kompleks antigen-antibodi terbentuk pada permukaan sel mast, dan yang terakhir menghasilkan reaksi alergi dengan membebaskan histamin, heparin, leukotrien dan ECF-A (faktor eosinofil-kemotaktik dari anafilaksis). Antigen semacam itu dikenal sebagai alergen.
5.IgD:
Itu ada sebagai monomer dan biasanya hadir dalam jejak sekitar 0,2% dari total imunoglobulin serum. Fungsi IgD tidak sepenuhnya dipahami. Karena IgD ( bersama-sama dengan IgM) ditemukan pada membran plasma limfosit-B, ia terlibat dalam diferensiasi sel-sel ini.
Distribusi Jaringan Limfoid:
- Folikel getah bening primer
- Kelenjar getah bening
- Haemolymph dan haemal node
- Timus
Folikel Limfe Primer (Gbr. 10-6):
Setiap folikel primer atau jaringan limfoid terdiri dari kumpulan limfosit Ð’ dan T yang didukung oleh serat retikuler. Bagian tengah folikel dikenal sebagai pusat germinal yang ditempati oleh limfoblas. Pinggiran folikel terdiri dari limfosit bebas dan sel plasma.
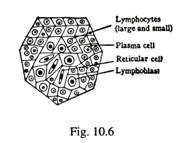
Folikel primer hadir dalam jaringan ikat longgar dari membran epitel basah saluran pernapasan bagian atas, saluran pencernaan dan saluran kemih. Mereka melawan masuknya antigen dari dunia luar. Folikel juga ada di kelenjar getah bening dan di limpa. Jaringan limfoid terkait mukosa (MALT) dalam kaitannya dengan usus dan bronkus masing-masing dikenal sebagai GALT dan BALT.
Keunikan dari folikel primer:
(a) Tidak adanya kapsul fibrosa yang pasti;
(b) Folikel menyaring cairan jaringan dan bertindak sebagai garis pertahanan kedua tubuh;
(c) Tidak memiliki pembuluh aferen, tetapi dilengkapi dengan pembuluh eferen.
Kelenjar getah bening:
Kelenjar getah bening biasanya tersusun dalam kelompok dan sering terletak di sepanjang pembuluh darah. Mereka bervariasi dalam bentuk dan ukuran. Rata-rata, setiap kelenjar getah bening berbentuk kacang dan memiliki hilus yang melekat pada satu pembuluh getah bening eferen. Sekitar 800 kelenjar getah bening ada di tubuh manusia.
Struktur Kelenjar Getah Bening (Gbr. 10-7):
Setiap node terdiri dari kapsul dan substansi kelenjar.
Kapsul fibrosa menutupi seluruh nodus dan dipisahkan dari substansi kelenjar oleh ruang subkapsular yang menerima terminal dari banyak pembuluh limfe aferen. Sejumlah trabekula meluas ke substansi kelenjar dari kapsul.
Ruang subkapsular dilintasi oleh serat retikuler kasar tempat sel retikuler melekat. Substansi kelenjar terdiri dari korteks luar dan medula dalam.
Korteks menyajikan hal-hal berikut:
(a) Banyak trabekula memanjang ke dalam dari kapsul dan membawa pembuluh darah. Setiap trabekula disertai oleh ruang para-trabekular yang terdiri dari serabut-serabut retikuler dan bersambung dengan ruang subkapsular.
(b) Daerah antara ruang paratrabekular ditempati oleh serat retikuler halus , celah yang diisi dengan sel-sel folikel limfe primer. Setiap folikel terdiri dari pusat germinal di tengah yang mengandung limfoblas, dan limfosit bebas serta sel plasma di pinggiran.
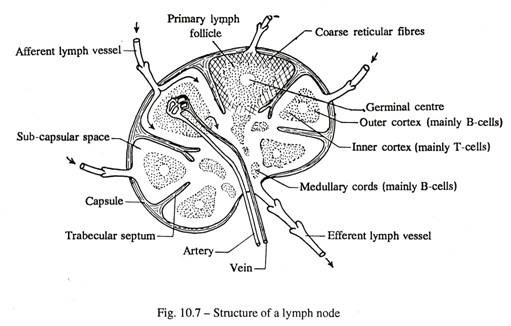
Di medula trabekula terbagi menjadi septa numerous. Ruang antara septa ditempati oleh tali limfosit yang tidak beraturan yang dikenal sebagai tali meduler. Akhirnya tali mencapai hilus kelenjar getah bening dari mana pembuluh getah bening eferen tunggal muncul.
Kerangka struktural kelenjar getah bening terdiri dari serat kapsul, trabekuler, dan retikuler. Celah serat retikuler diisi dengan sel retikuler tetap, dan limfosit bebas dan sel plasma.
Kekhasan kelenjar getah bening:
(a) Kehadiran kapsul berserat:
(b) Saring getah bening;
(c) Kehadiran pembuluh getah bening aferen dan eferen.
Fungsi Kelenjar Getah Bening:
(1) Kelenjar getah bening menyaring getah bening, dan menghilangkan partikel dan agen berbahaya (karbon, debu, bakteri, sel kanker) oleh aksi fagositik sel retikuler, saat getah bening meresap melalui ruang subkapsular dan para-trabekular . Virus, bagaimanapun, tidak disaring oleh node.
(2) Mereka menghasilkan limfosit yang dicuci keluar dari folikel getah bening ke dalam pembuluh getah bening eferen
Jumlah total limfosit yang dikirim oleh saluran toraks dalam 24 jam adalah sekitar 2,5 kali jumlah limfosit yang ada dalam aliran darah pada satu waktu. Pemeliharaan jumlah limfosit yang konstan dalam darah disediakan oleh resirkulasi limfosit (Gowans) dari kapiler darah trabekular ke ruang para-trabekular kelenjar getah bening, dan kemudian dicuci ke dalam pembuluh getah bening eferen (Gbr. 10- 7).
(3) Sel plasma dari folikel primer menghasilkan antibodi yang menghalangi aksi antigen bakteri atau agen asing lainnya.
(4) Limfosit B menetap di zona superfisial korteks dan di korda meduler kelenjar getah bening, sedangkan limfosit T biasanya terletak di zona tengah dan zona dalam korteks.
Haemolymph dan Haemal Nodes:
Nodus haemolymph terdiri dari campuran darah dan getah bening yang mengisi celah serat retikuler. Kelenjar ini jarang terjadi pada manusia, tetapi dapat ditemukan pada kelenjar getah bening retro-peritoneal.
Node Haemal:
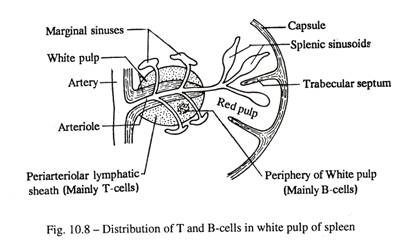
Limpa adalah simpul haemal, dan menyaring darah dengan mengeluarkan eritrosit, leukosit, trombosit dan antigen mikroba yang sudah usang dari sirkulasi. Terdiri dari kapsul, trabekula, serat retikuler, pulpa merah dan pulpa putih yang terbuat dari folikel limfe primer. Setiap folikel dilintasi secara eksentrik oleh arteriol. Limfosit T terletak di selubung limfe periarteriolar dan limfosit B menempati sisa pulpa putih (Gbr. 10-8).
timus:
Timus adalah struktur bilobed asimetris. Itu terletak di mediastina superior dan anterior toraks dan mengintervensi antara sternum di depan, dan perikardium, lengkung aorta dengan tiga cabangnya, vena brachio-cephalic dan trakea di belakang. Itu meluas ke bawah hingga kartilago kosta ke-4; di atas timus dapat meluas di depan trakea hingga kutub bawah lobus lateral kelenjar tiroid. Kedua lobus timus dihubungkan melintasi garis tengah oleh jaringan fibro areolar.
Saat lahir timus beratnya sekitar 10 gm sampai 15 gm; ukurannya semakin meningkat hingga usia pubertas ketika beratnya sekitar 20 gm hingga 30 gm. Setelah itu timus mengalami involusi dan diubah menjadi massa fibro-lemak; dalam kehidupan pertengahan dewasa beratnya mencapai sekitar 10 gram.
Setiap lobus ke timus berkembang dari endoderm kantong faring ketiga dan mengalami migrasi kaudal di toraks. Lampiran dasar timus ke faring primitif kemudian terputus . Sel-sel endodermal bertahan sebagai tali sel epitel retikuler.
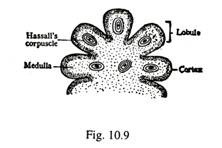
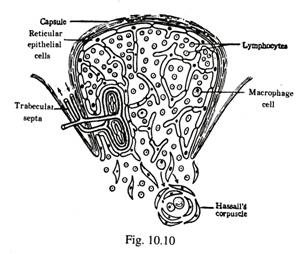
Struktur Timus (Gbr. 10-9, 10-10):
Setiap lobus timus ditutupi oleh kapsul fibrosa yang menonjol ke dalam substansi organ sebagai septa trabekular yang tidak lengkap. Septa trabekular membawa pembuluh darah dan membagi timus menjadi banyak lobulus; setiap lobulus memiliki lebar sekitar 1 mm hingga 2 mm. Lobulus terdiri dari korteks luar dan medula dalam. Korteks mengandung limfosit numerous yang padat dan kadang-kadang sel makrofag.
Di medula jumlah limfosit lebih sedikit; selain itu berisi sel darah Hassall yang konsentris. Beberapa pembuluh darah dari septa trabekular melintasi persimpangan antara korteks dan medula, dan terbagi menjadi kapiler kortial dan meduler. Timus tidak memiliki kapiler getah bening.
Beberapa Detail tentang Struktur:
Empat jenis struktur khusus ditemui di timus. Ini adalah sel epitel retikuler, limfosit, makrofag, dan sel darah Hassal.
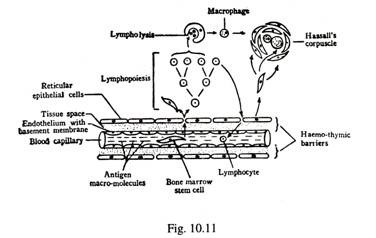
Sel-sel epitel retikular dari lembaran kontinu yang melapisi permukaan bagian dalam kapsul fibrosa, trabecular septa, dan sekitar pembuluh darah di persimpangan kortiko-medula dan kapiler kortikal dan meduler. Sel-sel retikuler dihubungkan satu sama lain oleh demosom. Kabel bercabang tidak teratur dari sel epitel retikuler bercabang di korteks dan medula terlebih lagi di bekas. Celah antara sel retikuler diisi dengan banyak limfosit dan kadang-kadang makrofag. Makromolekul antigen dari darah yang bersirkulasi dicegah untuk bersentuhan dengan limfosit timus karena adanya Haemo-Thymic Barrier.
Penghalang terdiri dari yang berikut dari luar ke dalam — lapisan sel endotel kapiler yang terus menerus; bran membran basement yang tebal ; ruang jaringan yang kadang-kadang berisi cairan jaringan; lapisan sel epitel retikuler yang kontinu (Gbr. 10-11). Meskipun penghalang tidak permeabel terhadap antigen, zat nutrisi dan sel punca dari sumsum tulang diangkut ke timus melalui penghalang. Selain itu, limfosit thymus melewati penghalang ke kolam sirkulasi.
Limfosit dalam timus berasal dari sel punca sumsum tulang. Limfosit berkembang biak secara asimetris dalam lingkungan bebas antigen. Beberapa sel diawetkan sebagai sel induk untuk pembelahan sel selanjutnya , sedangkan sel lainnya berkembang biak berulang kali dengan mitosis untuk membentuk banyak limfosit kecil.
Sel-sel ini mengisi lebih tebal di korteks daripada medula, dan mereka menempati celah antara sel epitel retikuler. 90% berat timus disumbangkan oleh limfosit. Sebagian besar limfosit timus (90%) berumur pendek dengan rentang hidup 3 sampai 5 hari. Mungkin sel-sel ini menjadi alergen otomatis satu sama lain atau ke inang, dan mengalami disintegrasi dini. Limfosit yang mengalami degenerasi difagositosis oleh makrofag. Sekitar 5% dari limfosit yang masih hidup muncul di kolam sirkulasi melalui penghalang hemotimik sebagai sel yang kompeten secara imunologis, dan bersirkulasi ulang di folikel limfatik primer dari organ perifer sistem limfatik.
Timus bertindak sebagai organ sentral dari sistem limfatik dan menyediakan limfosit yang tidak terikat yang bereaksi terhadap berbagai antigen baru, sedangkan limfosit organ perifer berkomitmen untuk bereaksi dengan antigen spesifik. Selain itu, timus mengatur proliferasi limfosit baik di dalam timus maupun di organ limfa perifer, misalnya kelenjar getah bening.
Zona tengah dan dalam kelenjar getah bening dianggap sebagai area yang bergantung pada timus. Limfopoiesis di dalam timus dan di organ perifer mungkin diatur oleh faktor humoral, limfopoietin, yang dipisahkan oleh sel epitel retikuler timus. Limfopoiesis timus dan limfolisis bersifat otonom, dan hanya dikendalikan oleh timus.
Sel epitel retikuler medula lebih eosinofilik. Beberapa dari sel-sel ini mengalami pembengkakan dan nukleusnya terfragmentasi. Sel-sel yang hancur tersebut membentuk massa hyalinzing sentral secara individual. Setiap massa pusat dikelilingi secara konsentris oleh lapisan sel epitel eosiÂnofilik dan membentuk sel darah Hassall. Sel-sel makrofag yang mengandung limfosit yang difagosit dimasukkan ke dalam sel-sel darah Hassal yang konsentris. Sel-sel ini berdiameter sekitar 30 hingga 100 µm dan jumlahnya banyak selama involusi timus.
Pengaruh Hormon pada Timus:
Hormon pertumbuhan hipofisis anterior dan hormon tiroid merangsang timus untuk tumbuh sebelum mencapai pubertas. Kehadiran hormon steriod dari korteks suprarental dan dari gonad mendukung involusi timus, yang biasanya diamati pada kehidupan pasca pubertas.
Pengebirian atau adrenalektomi pada awal kehidupan menunda involusi timus. Di sisi lain, pemberian kortison (hormon kortikal suprarenal) menghasilkan involusi awal timus dan menekan limfopoiesis dari seluruh sistem limfatik.
Fungsi Timus:
Dalam beberapa tahun terakhir, studi tentang thymus mengungkapkan beberapa fungsi menarik dan berguna, yang sampai sekarang belum dijelajahi.
(1) Timus bertindak sebagai organ sentral dari sistem limfatik dan memberikan limfosit yang kompeten secara imunologis ke kolam sirkulasi dan ke organ limfatik perifer. Limfosit timus (limfosit-T) tumbuh di lingkungan bebas antigen dan menghasilkan sel-sel bebas yang dapat bereaksi dengan varietas antigen baru.
(2) Sangat penting dalam minggu-minggu awal kehidupan neonatal dan mengatur pertumbuhan jaringan limfoid perifer. Pemberian kortison pada bayi baru lahir merugikan karena menghambat respon imunologi normal.
(3) Timus membesar pada beberapa penyakit autoimun, misalnya myesthenia gravis. Pada myesthenia gravis, otot-otot volunter tertentu mengalami kelelahan dini setelah beberapa kontraksi awal. ThymecÂtomy dalam kondisi ini memperbaiki gejala. Mungkin timus membebaskan beberapa zat penghambat seperti curare yang menghalangi transmisi neuromuskular.
Sel dalam Respon Kekebalan Tubuh:
Untuk melawan invasi antigen yang sangat besar, tubuh dilengkapi dengan tiga kelompok sel untuk pertahanan diri – limfosit Ð’ (sel B), limfosit T (sel T) dan sel penyaji Antigen (APC)
Ketika tidak aktif, baik sel Ð’ dan sel T adalah limfosit kecil dengan diameter 6-10 pm, dan masing-masing memiliki nukleus bulat dan pinggiran tipis sitoplasma yang sedikit di sekitar nukleus. Tetapi ketika terkena antigen tertentu, sel-sel tersebut teraktivasi membentuk limfosit besar dan berdiferensiasi menjadi limfosit efektor Ð’ dan T. Baik sel Ð’ dan T bervariasi dalam rentang hidup; beberapa hanya hidup beberapa hari, sementara yang lain bertahan dalam sirkulasi darah selama bertahun-tahun (sel memori).
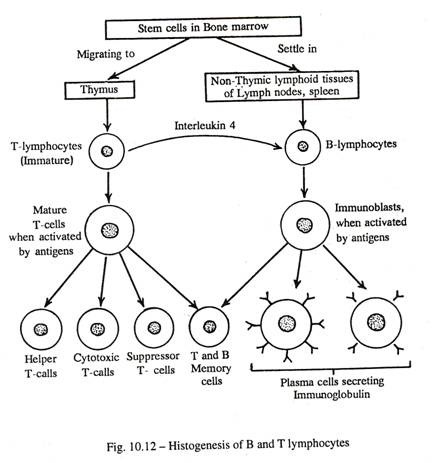
limfosit Ð’ (Gbr. 10-12 dan 10-13):
Pada mamalia, secara umum diyakini bahwa prekursor sel Ð’ diproses di sumsum tulang, di mana mereka berdiferensiasi dalam lingkungan mikro khusus menjadi limfosit Ð’ yang matang atau efektor setelah mitosis berulang. Namun, pada burung, sel Ð’ berasal dari kantong endodermal, bursa Fabricius, yang melekat pada usus belakang; oleh karena itu dinamakan limfosit B atau limfosit setara bursa. Tetapi keberadaan bursa kloaka seperti itu diragukan pada mamalia.
Sel-sel Ð’ dewasa atau efektor meninggalkan sumsum tulang dan menetap di: (a) kelenjar getah bening di dalam zona superfisial korteks; (b) dalam pulpa putih limpa di luar selubung limfatik periarteriolar ; (c) dalam jaringan limfoid difus di bawah selaput lendir sistem pernapasan, pencernaan dan kemih; (d) beberapa sel bebas bersirkulasi dalam darah. Sekitar 20% limfosit yang bersirkulasi merupakan sel Ð’.
Ketika diaktifkan oleh antigen spesifik, sel efektor Ð’ berproliferasi melalui mitosis, dan berdiferensiasi menjadi sel plasma yang mensekresikan sirkulasi antibodi atau imunoglobulin, yang spesifik antigen. Jadi sel Ð’, sel plasma dan antibodi membentuk dasar imunitas humoral (vide supra). Diferensiasi sel Ð’ menjadi sel plasma dibantu oleh limfokin yang disekresikan oleh limfosit T pembantu dan oleh antibodi IgM dan IgD yang terikat membran. Beberapa sel yang diaktifkan bertahan sebagai sel Ð’ memori yang telah diprogram sebelumnya yang akan menghasilkan respons imun cepat pada paparan berikutnya oleh antigen spesifik yang sama.
Sel Ð’ mengenali antigen melalui kompleks reseptor antigen. IgM, hadir pada permukaan semua sel Ð’, merupakan komponen pengikat antigen dari reseptor sel Ð’. Beberapa molekul lain pada permukaan sel-B sangat penting untuk fungsi sel Ð’. Ini termasuk reseptor pelengkap, reseptor Fc dan CD40. Molekul CD40 memainkan peran penting dalam interaksi sel T helper dan sel B. Interaksi ini sangat penting untuk pematangan sel Ð’ dan sekresi IgG, IgA, dan IgE.
T-limfosit (Lihat Gambar 10-12, 13):
Sel induk primitif limfosit T diturunkan dari sumsum tulang, dan meninggalkan ruang sumsum melalui sistem peredaran darah untuk muncul di timus, di mana sel T mengembangkan kematangan imunokompeten dengan
mitosis berulang dalam lingkungan bebas antigen karena adanya barier hemotimik. Kematangan sel T dibantu oleh sel retikuler timus dan makrofag. Dalam proses ini banyak limfosit yang bermusuhan dengan antigen diri dikenali, dihancurkan atau ditekan oleh mekanisme yang tidak diketahui.
Sel T yang bertahan hidup berkembang menjadi sel imunokompeten untuk antigen nondiri, dibebaskan dalam sirkulasi dan menetap di area berikut: (a) kelenjar getah bening di zona tengah korteks (zona parakortikal); (b) selubung limfatik periarteriolar limpa; (c) jaringan limfoid difus dari sistem limfatik mukosa; (d) limfosit bebas dalam sirkulasi darah. Sekitar 75% limfosit yang bersirkulasi berasal dari sel T.
Ketika distimulasi oleh antigen, beberapa sel T mengeluarkan limfokin yang semuanya adalah peptida atau protein dan mempengaruhi pertumbuhan dan diferensiasi antara berbagai kelompok sel imun. Limfokin juga dikenal sebagai Interleukin.es (IL); lebih dari enam belas interleukin telah dijelaskan sejauh ini. Dari jumlah tersebut, IL 4 yang disekresikan oleh sel T merangsang diferensiasi sel Ð’.
Semua sel T memiliki protein reseptor sel T (TCR) di permukaan, yang mengenali antigen spesifik yang mirip dengan antibodi. Karena sel T berfungsi dengan mengarahkan dan merekrut sel lain tanpa mensekresikan anti -tubuh, mereka membentuk dasar imunitas seluler. Sekitar 5% limfosit dalam darah disebut sel nol. Mereka tidak memiliki antigen permukaan limfosit T atau Ð’, dan diyakini sebagai sel punca yang bersirkulasi.
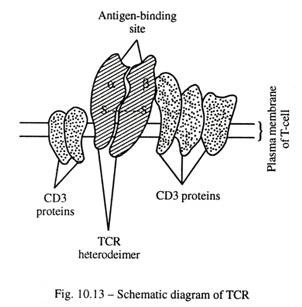
reseptor sel-T:
TCR terdiri dari heterodimer terkait disulfida, rantai polipeptida a dan ap di sebagian besar sel T, masing-masing memiliki variabel (pengikatan tigen) dan wilayah konstan. Dalam minoritas sel T, TCR terdiri dari у dan 5 rantai polipeptida; seperti (γ/δ) sel TCR cenderung berkumpul di permukaan epitel saluran pernapasan dan GI.
Kedua jenis TCR di atas terkait dengan sekelompok lima rantai polipeptida, yang dikenal sebagai kompleks molekuler CD3. Mereka terlibat dalam transduksi sinyal ke sel-T setelah mengikat antigen (Gbr. 10-12).
Kelas sel T:
Dengan penerapan antibodi monoklonal terhadap limfosit, sekarang dimungkinkan untuk mengklasifikasikan limfosit T dengan cluster of differentiaion (CD) sebagai molekul penanda pada permukaan sel. Semua sel T sejati adalah CD 3 positif; selain itu CD 4 positif terhadap sel T penolong, dan CD 8 positif terhadap sel T sitotoksik dan penekan. Sel pembunuh alami adalah CD 3 positif, tetapi tidak mengandung penanda CD 4 dan CD 8 . Dengan demikian, sel T terdiri dari empat subtipe – sel T pembantu, sitotoksik, penekan dan memori.
Pengenalan antigen oleh sel-T:
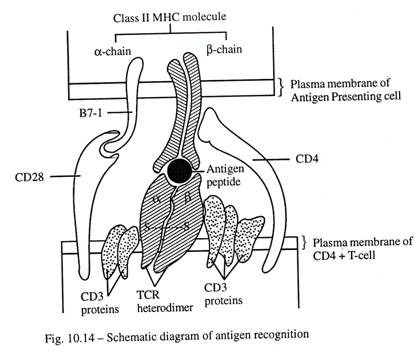
Selain protein CD3, sel-T mengekspresikan berbagai molekul yang terkait secara fungsional di permukaan sel yang meliputi CD4, CD8, CD28, CD40, dan banyak lainnya. Sekitar 60% sel T dewasa adalah CD4+ dan 30% CD8+. Selama aktivasi sel T, molekul CD4 dari sel T penolong bertindak sebagai koreseptor dan berikatan dengan molekul kompleks histokompatibilitas mayor (MHC) kelas II pada sel penyaji antigen, sedangkan sel T sitotoksik CD8+ mengenali ikatan sel antigen hanya berasosiasi dengan molekul MHC kelas I.
Tetapi untuk aktivasi sel T CD4+ dan CD8+ yang berkepanjangan, diperlukan interaksi antara molekul CD28 pada sel T dan molekul B7-1 atau B7-2 yang diekspresikan pada sel penyaji antigen . Jika tidak, sel-T akan hancur atau menjadi tidak reaktif (Gbr. 10-14).
Helper T Cells ( TH ): Ini memainkan peran yang sangat penting dari imunitas seluler dengan mengeluarkan limfokin, ketika diaktifkan oleh kombinasi molekul antigen dan kelas II MHC (major histocompatibility complex) yang terikat pada mambrane plasma sel makrofag. Baru-baru ini dua himpunan bagian dari sel-T helper dikenali:
1) Subset TH -1 mengeluarkan interleukin-2 (IL-2) dan interferon-y (IFN-γ) . Ini terlibat dalam memfasilitasi hipersensitivitas tertunda, aktivasi makrofag dan sintesis antibodi IgG-2b.
2) Subset TH -2 menghasilkan IL-4 dan IL-5. Ini membantu dalam sintesis kelas antibodi lainnya.
Limfokin, juga disebut sitokin, melakukan fungsi-fungsi berikut:
saya. Merangsang proliferasi dan pematangan sel T sitotoksik dan penekan;
- Interleukin 4, salah satu jenis lympho kines, mengaktifkan klon sel В yang belum matang untuk menghasilkan antibodi melalui sel plasma terhadap antigen tertentu tersebut. Jadi sel-sel TH meningkatkan serangan terhadap antigen itu dengan imunitas seluler dan humorial .
aku aku aku. γ-interferon, disekresikan oleh sel TH, menginduksi ekspresi Kelas IIMHC dan mengaktifkan makrofag .
Penanda utama sel T adalah CD, molekul; karenanya disebut sel T4 .
Kepentingan klinis: Sel T pembantu dibunuh oleh virus HIV yang menyebabkan Acquired Immune Deficiency Syndrome, AIDS. Akibatnya, kekebalan pasien yang terinfeksi lumpuh dan membuat mereka rentan terhadap infeksi oportunistik.
Sel T sitotoksik (Tc ) :
Sel-sel ini membunuh sel-sel yang terinfeksi virus dan ganas, dan sel-sel ‘al ien’ dari allograft. Hal ini dilakukan dengan pelepasan protein lisosomal beracun, perforin, yang menghasilkan lubang pada membran sel dari sel target.
Protein reseptor sel Tc mengenali sel patologis atau sel cangkok yang berbeda secara genetik dengan menggabungkan antigen asing dalam kombinasi dengan molekul MHC kelas I yang menempel pada membran sel sel target. Tc celt diaktifkan oleh limfokin sel T H.
Sel Pembunuh Alami (sel NK):
Morfologi dan aksinya hampir mirip dengan sel Tc, tetapi tidak mengandung penanda CD 4 atau CD S. Sel NK diaktifkan oleh Interleukin 2 dari sel TH . Mereka membebaskan cytolysin dan membunuh sel yang tidak terinfeksi virus secara khusus, protozoa dan sel patogen lainnya.
Sel T Penekan (Ts):
Sel-sel ini menghambat fungsi sel TH dan Tc , dan memodulasi antara kontrol positif dan negatif dari respons imun. Hal ini menjadi keharusan karena penghancuran penyerang asing yang berlebihan oleh agen kuat dapat melibatkan tubuh itu sendiri dalam bentuk penyakit autoimun, asma alergi dan dermatitis alergi.
CD S berperan sebagai penanda sel T supresor.
Sel Memori T: Pada paparan pertama terhadap antigen, beberapa klon sel T yang tidak aktif mengembangkan protein permukaan reseptor yang spesifik untuk gen anti itu, tetapi mereka sendiri tidak terlibat dalam respons imun. Ini bertahan sebagai sel T memori, yang memiliki rentang hidup yang panjang. Saat terpapar antigen yang sama pada kesempatan berikutnya, respons sel T memori segera menantang penyerbu.
Sel penyaji antigen (sel APC):
APC ditemukan di sebagian besar jaringan. Mereka memfagositosis antigen, memprosesnya dan mempertahankan produknya untuk waktu yang lebih lama dan secara bertahap menghadirkannya ke permukaan sel dalam kombinasi dengan molekul protein MHC dan mengaktifkan limfosit (Gbr. 10-14).
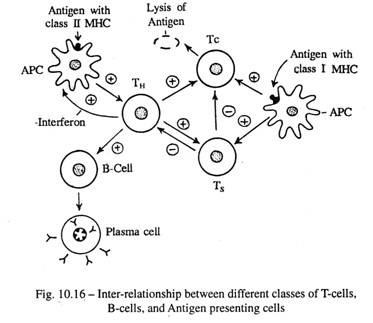
APC merupakan populasi sel heterogen yang termasuk dalam sistem fagosit mononuklear (MPS). Mereka termasuk makrofag, sel Langerhans epidermal, sel dendritik organ limfoid, sel epitel retikuler timus, dan mikroglia sistem saraf pusat.
Diproses oleh APC:
Antigen dibawa ke APC dan sebagian terfragmentasi. Ini kemudian terikat pada salah satu kelas antigen histokompatibilitas yang dikenal sebagai protein MHC dan seluruh kompleks molekul terpapar ke permukaan sel dari sel penyaji antigen.
Ketika antigen yang diproses bertemu dengan sel T matang yang mengandung reseptor yang sesuai, ia mengaktifkan sel T. Ketika antigen yang diproses terikat pada protein MHC kelas I, ia menghasilkan respons sel Tc atau sel Ts yang mengandung CD K. Di hadapan protein MHC kelas II yang terikat pada gen antigen yang diproses, ini mengaktifkan respons sel TH , yang pada gilirannya membantu sel Ð’ untuk merespons antigen yang sama.



