Artikel ini memberikan informasi tentang ultrastruktur mitokondria, jenisnya, butiran terkait dan partikel mitokondria!
Mitokondria terdiri dari dua bagian:
(1) membran pembatas, dan
(2) massa dalam atau matriks Palade, (1952).
1. Membatasi membran:
Membran ini ganda. Membran luar memiliki ketebalan sekitar 60 hingga 75 A. Membran bagian dalam tebalnya sekitar 50 sampai 70 A dan menunjukkan banyak lipatan (seperti pelat) ke dalam rongga mitokondria, yang disebut puncak atau krista mitokondria. Mereka menembus matriks mitokondria.
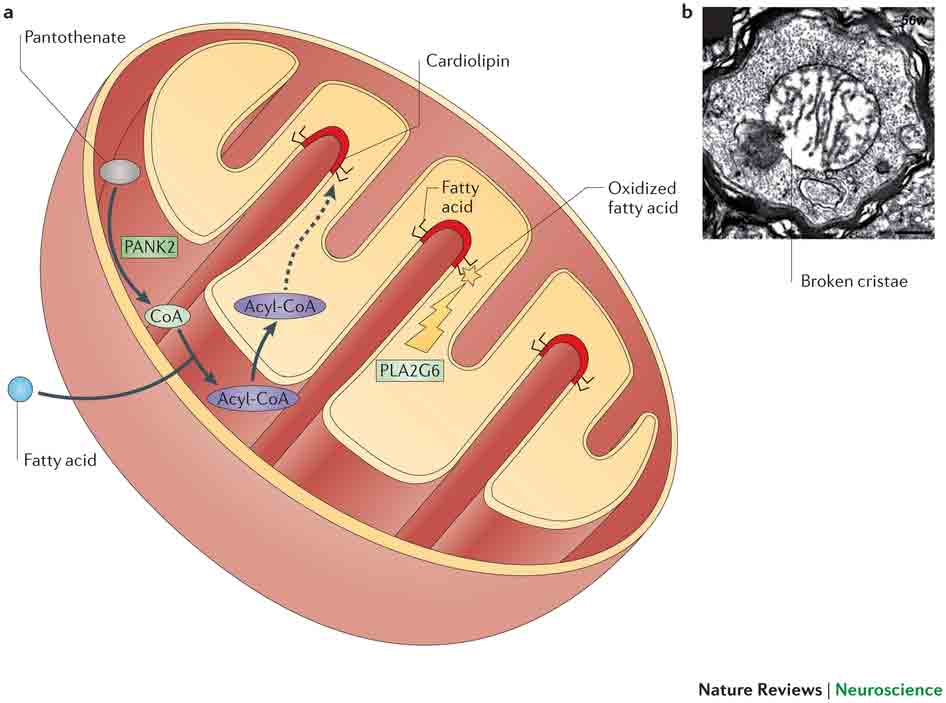
Sumber Gambar : nature.com/nrn/journal/v14/n8/images/nrn3453-f4.jpg
Krista (krista singular) sangat memperluas area permukaan yang terpapar ke rongga bagian dalam mitokondria, menyediakan ruang yang cukup untuk akomodasi enzim. Krista biasanya berjalan di sudut kanan ke sumbu panjang mitokondria berbentuk batang.
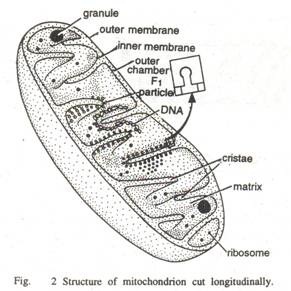
Ruang antara membran luar dan dalam disebut ruang perimitochondrial yang tebalnya 60 sampai 80 A (Tylers, 1973), dan kurang padat. Itu juga meluas di puncak. Masing-masing membran ini terdiri dari lapisan protein luar, lapisan bimolekuler tengah lipid dan lapisan protein dalam, yaitu membran memiliki struktur trilaminar. Lapisan protein osmofilik padat luar dan dalam setebal 20 hingga 25 A.
2. Matriks:
Itu diisi dengan bahan cairan protein yang relatif padat yang umumnya homogen, tetapi dalam beberapa kasus mungkin mengandung bahan berfilamen halus atau butiran kecil yang sangat padat dengan diameter sekitar 40 hingga 50 A yang dikenal sebagai butiran intra mitokondria.
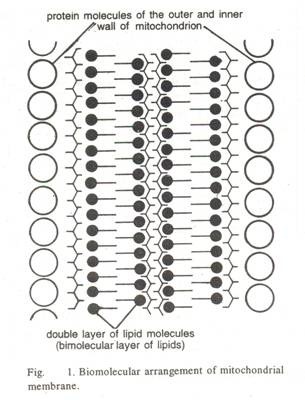
Di dalam matriks terdapat ribosom kecil, RNA dan satu atau lebih molekul DNA sirkular, dengan panjang sekitar 4,7 hingga 5,4 µ. Ini memiliki banyak hasil sebagai hasil dari lipatan membran dalam. Lipatan ke dalam ini disebut sebagai krista. Ribosom lebih kecil dari ribosom eukariotik sitoplasma dan lebih mendekati ukuran ribosom prokariotik, berukuran diameter sekitar 120-150 A dan RNA transfer.
Karakteristik butiran matriks adi-elektronik (padat) rata-rata berdiameter 300 A. Butiran ini sangat sedikit atau tidak ada dalam mitokondria Protozoa, sel miokard dan sel tiroid istirahat; sedangkan dalam sel penyerap duodenum, hati, pankreas, dan sel perbatasan sikat, butiran о sangat banyak.
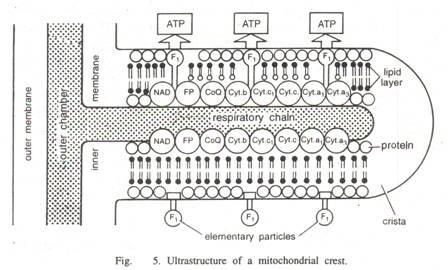
Granula ini berfungsi sebagai tempat pengikatan ion kation divalen magnesium dan kalsium (Ca ++ dan Mg ++ ), yang berperan penting dalam enzim. Membran bagian dalam dan krista dapat dilihat terdiri dari partikel elementer (atau rakitan pernapasan), masing-masing terdiri dari dasar dan kepala bulat (berdiameter 8 hingga 9 mm) yang dihubungkan oleh bagian penghubung (atau tangkai). Partikel dasar mengandung enzim yang terlibat secara khusus dalam pembentukan ATP.
Matriks berisi semua enzim larut yang terlibat dalam siklus Krebs, selain sintesis DNA dan protein. Membran bagian dalam membawa semua enzim yang terkait dengan rantai pernapasan (ATPase) dan fosforilasi, serta protein pembawa spesifik yang terlibat dalam perembesan metabolit seperti ADP, ATP, dan fosfat. Susunan krista bervariasi dan mungkin sebagai berikut —
(1) Mereka mungkin sejajar dengan sumbu panjang mitokondria, seperti pada neuron dan sel otot lurik.
(2) Umumnya mereka terletak tegak lurus terhadap sumbu panjang.
(3) Ini mungkin dalam bentuk vesikel, sering bercabang untuk membentuk jaringan ruang penghubung, seperti pada sel kelenjar paratiroid, leukosit manusia, beberapa protozoa, dll.
(4) Krista ini juga ditemukan dalam susunan tubular pada sel kelenjar adrenal dan telah ditemukan pada sel meristematik tanaman dan tubulus Malphigi pada serangga.
(5) Pada spermatid tertentu, krista tersusun secara konsentris di dalam matriks.
(6) Pada Amoeba banyak krista saling bertautan membentuk vili.
Variasi jumlah krista berhubungan dengan volume matriks. Di mana krista relatif sedikit (misalnya, hati mamalia), terdapat banyak matriks, dan sejumlah besar krista (jaringan otot) mengurangi jumlah matriks.
Jumlah krista secara langsung mempengaruhi kapasitas mitokondria untuk melakukan reaksi oksidatif. Dalam sel otot terbang serangga, terdapat lebih banyak krista dan juga lebih banyak mitokondria daripada sel tubuh lainnya.

[I] Jenis-jenis krista:
Cristae dapat dibagi menjadi dua jenis: (1) Septate dan (2) Tubular.
1. Septate (lengkap atau tidak lengkap) cristae:
Ini adalah partisi bersisi paralel yang muncul sebagai tiga lapis. Partisi (septa) terdiri dari dua unit membran yang dipisahkan oleh kelanjutan ruang luar. Biasanya, septate cristae tunggal dan lurus; tetapi pada mukosa lambung katak dan pankreas kelelawar, mereka membentuk angulasi, yaitu krista tidak tersusun dalam barisan paralel tetapi menyimpang pada sudut 90° terhadap yang lain yang terkadang menghasilkan pola zig-zag. Krista septate terkait adalah tipe prismatik, terjadi pada otot krikotiroid laring kelelawar.
Di dalamnya, krista tersusun rapat, berorientasi melintang dan juga berjendela. Mereka membentuk profil segitiga dengan panjang 600 A yang memiliki tubulus prismatik, yang muncul sebagai strip paralel jika dipotong secara longitudinal. Akhirnya, septate cristae dapat membentuk partisi kompleks selain yang sederhana. Di dalamnya, krista dapat bercabang; interkoneksi untuk menimbulkan pola sisir madu seperti pada leukosit manusia.
2. Tubular (villous) cristae:
Ini muncul sebagai partisi membran dalam yang mirip vili, ditemukan di mitokondria protozoa, sel hati, dan neuron. Bagian terminal tubulus dapat melebar membentuk area bulat dan krista ini disebut krista sakular atau vesikuler.
Pada amoeba raksasa, Pelomyxa carolinensis, krista tubular tersusun dengan cara tertentu. Di salah satu ujung mitokondria, krista sedikit dan berbentuk tidak teratur tetapi ujung lainnya mengandung krista yang tersusun dalam 3 jenis pola zig-zag.
Lipatan atau krista ini memberikan peningkatan luas permukaan dalam mitokondria untuk aktivitas enzimatik.
Krista memiliki partikel F 1 dan faktor kopling. Selanjutnya aktivitas beberapa enzim, (yakni suksinat dehidrogenase, β-hidroksi butir dehidrogenase, rotenone sensitif NADH-sitokrom-C-reduktase dan ATPase) mendominasi dalam krista.
[II] Butiran terkait:
Pada beberapa protozoa dan jamur tertentu, permukaan dalam membran mitokondria menjadi terkait dengan butiran kecil. Sel-sel pankreas babi guinea menunjukkan peningkatan jumlah butiran ini dengan puasa, terutama karena mitokondria menjadi terkait dengan tetesan lipid. Mereka mungkin terlibat dalam aktivitas metabolisme mitokondria.
[III] Partikel mitokondria atau partikel Fi atau partikel elementer:
Mikroskop elektron telah mengungkap keberadaan partikel yang sangat kecil di dalam membran dalam dan di luar membran luar. Mereka pertama kali dijelaskan oleh Humberto Fernandez-Moran (1963). Partikel membran luar tidak bertangkai, muncul sebagai bola sederhana.
Ini diyakini melakukan reaksi oksidasi yang memasok elektron dan juga berfungsi sebagai situs untuk mengkatalisasi reaksi sintetik dengan ATP. Setiap partikel bertangkai dari membran dalam, disebut partikel elementer atau partikel F1, terdiri dari basis berbentuk kubus 40 × 110 A, batang berdiameter 30 – 40 A dan panjang 45-50 A, dan kepala bundar 75 hingga 100 A in diameter.
Ini terkait erat pada permukaan krista dan secara teratur ditempatkan pada jarak 100 A. Tergantung pada ukuran dan jenis mitokondria, terdapat 10.000 hingga 100.000 partikel elementer per mitokondria.
Secara fungsional partikel elementer berhubungan dengan keberadaan enzim untuk fosforilasi oksidatif dan dengan mitokondria adenosin trifosfatase. Oleh karena itu partikel-partikel ini disebut partikel transpor elektron atau ETP (Parsons, 1963). Namun baru-baru ini Racker (1967) telah menunjukkan bahwa partikel F1 (ETP) mewakili ATPase atau ATP sintetase khusus dan mengambil bagian dalam proses fosforilasi oksidatif.
ETP mengkatalisis oksidasi NADH dan suksinat tetapi tanpa fosforilasi berpasangan (Crane, et al., 1956). Asli mengandung beberapa protein struktural dan sekitar 30% lipid. Itu telah dipisahkan menjadi empat kompleks yang sejauh ini merupakan unit operasional terkecil yang diisolasi dari mitokondria –
Kompleks I:
Mengkatalisis oksidasi NADH digabungkan dengan reduksi koenzim Q, dan mengandung 1 mol flavin (mungkin sebagai NADH dehydrogenase), 20% lipid, 3 mol koenzim Q (larut lipid), dan 16 atom besi nonheme.
Kompleks II:
Mirip dengan kompleks I, tetapi mengkatalisasi oksidasi suksinat yang digabungkan dengan reduksi koenzim Q. Ini mengandung 1 mol flavin (sebagai suksinat dehidrogenase), mol I sitokrom b, lipid, koenzim Q dan besi nonheme.
Kompleks III:
Mengkatalisis oksidasi koenzim Q digabungkan dengan reduksi sitokrom C. Kompleks ini berwarna merah dan mengandung dua mol sitokrom b, satu mol sitokrom C 1 , satu mol protein besi nonheme, dan lipid.
Kompleks IV:
Mengandung sitokrom a dan а 3 (sitokrom oksidase) dan berwarna hijau.
Berbagai kompleks ini dapat bergabung kembali satu sama lain dalam rasio 1: 1: 1: 1 membangun kembali seluruh rantai transfer elektron dari NADH atau suksinat ke oksigen. Namun, karena koenzim Q dan sitokrom С tidak terikat secara struktural, mereka harus ditambahkan kembali ke persiapan oksidasi yang efektif. Green telah menyarankan bahwa setiap kompleks melakukan fosforilasi berpasangan.
Rantai Pernafasan atau Sistem Transportasi Elektron (ETS):
Sistem transpor elektron terdiri dari serangkaian protein kompleks, yang mengambil bagian dalam rantai pernapasan. Ada empat kompleks yang terbentuk dari lipoprotein dan dua pembawa elektron bergerak-koenzim Q (CoQ) atau ubiquinone (UQ) dan sitokrom C.
[I] Kompleks:
Kompleks adalah situs di mana ion hidrogen yang dilepaskan selama siklus kreb dioksidasi dan energinya terperangkap dalam ATP.
Menurut pandangan saat ini (Hinkle dan McCarty, 1978) potongan kepala partikel mitokondria mengandung ATPase yang tepat, tangkai terdiri dari F 5 atau oligomycin-sensitivity conferring protein (OSCP) dan Fe (F C2 ) dan bagian dasar (F O ) memiliki saluran protein. Singkatnya, struktur dan komponen rantai pernapasan diberikan di bawah ini –
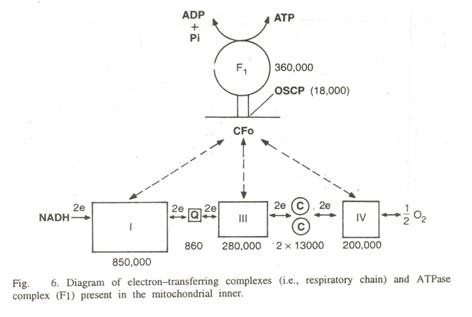
- Kompleks I:
(NADH—CoQ reduktase). Ini memiliki komponen berikut:
(a) NADH dehidrogenase:
Ini terdiri dari flavoprotein dengan FMN sebagai kelompok prostetik. Proteinnya adalah rantai polipeptida tunggal dengan mw 70.000.
(b) Besi nonheme (NHI):
Protein dengan pusat besi-sulfur (Fe-S). Ada enam pusat Fe-S, yaitu Fe-SNla, Fe-SNlb, Fe-SN2, Fe-SN3, Fe-SN4 dan Fe-SN5. Total mw 850.000. Ini adalah kompleks terbesar dan termasuk flavoprotein yang mengandung FMN. Ini adalah langkah pertama dalam rantai transpor elektron. Elektron dibawa ke kompleks ini oleh NAD + yang terletak di sisi matriks membran.
- Kompleks II:
(Suksinat—CoQ reduktase). Ini terdiri dari komponen-komponen berikut –
(i) Suksinat dehidrogenase dengan mw 70.000 memiliki FAD yang terikat secara kovalen sebagai gugus prostetik dan dua pusat Fe-S, yaitu Fe-SSl dan Fe-SS2.
(ii) protein Fe-SS 3 mw 27.000 dan
(iii) Sitokrom b dengan absorbansi 557,5 nm.
Koenzim Q:
(CoQ) atau Ubiquinone (UQ). Ini adalah pembawa seluler antara kompleks I dan III, dan II dan III. Kompleks II mendahului rantai transpor elektron dan digabungkan menjadi suksinat melalui FAD (flavin adenine dinucleotide).
- Kompleks III:
(CoQH2-Cyt.C-reduktase). Kompleks ini mengandung —
(i) Sitokrom b dari mw 30.000
(ii) Sitokrom e dari mw 50.000
(iii) Sitokrom С 1 memiliki dua polipeptida mw 29.000 dan 15.000.
(iv) protein NH 1 dengan pusat FeS dan mw 26.000.
(v) Protein inti
(vi) Protein pengikat antimisin.
Sitokrom c:
Ini adalah pembawa bergerak antara kompleks III dan IV dengan mw 13.000. Di dalamnya terdapat satu heme Ñ yang terikat pada rantai polipeptida.
- Kompleks IV:
(Sitokrom c-oksidase). Ini mengandung sitokrom a (Cyt. a) tidak dihambat oleh CO, sitokrom аз (Cyt. аз) dihambat oleh CO dan dua atom tembaga (Cu dan Cu). Ini adalah oksidasi akhir hidrogen, menghasilkan air (H2O).
- Kompleks V (kompleks ATPase):
Ini berisi bagian kepala, tangkai dan bagian dasar. Bagian kepala (F 1 ) terdiri dari 5 subunit dan inhibitor mw 3, 60.000.
α — Subunit 2 atau 3 dengan mw 53.000
β— Subunit 2 atau 3 dengan mw 50.000
γ— Subunit 1 atau 2 dengan mw 33.000
δ – Subunit 1 atau 2 dengan mw 17.500
E— Subunit 1 atau 2 dengan mw 7000
F 1 inhibitor protein I dengan mw 10.000
Tangkai memiliki F 5 atau oligomycin sensitivity conferring protein (OSCP) sebesar mw 18.000 dan F6 (Fe 2 ) sebesar mw 8.000.
Bagian dasar (Fo) terbuat dari proteolipid—kompleks protein hidrofobik yang membentuk saluran proton. Ada 4 protein mw 29.000, 22.000, 12.000 dan 7800.
Semua kompleks ini dan sistem fosforilasi diatur dalam membran mitokondria bagian dalam dalam susunan yang sangat asimetris. Sistem transpor elektron hanya dapat diakses oleh NADH dan suksinat dari sisi matriks membran, sedangkan sitokrom Ñ dicapai dari sisi sitoplasma membran. Organisasi molekuler ini konsisten dengan transfer proton (H + ) melintasi membran dari sisi matriks ke sisi sitoplasma membran.
Rantai pernapasan digabungkan pada tiga titik dengan sistem di mana terjadi fosforilasi ADP menjadi ATP. Keenam proton yang berasal dari rantai pernapasan ditranslokasi melintasi membran mitokondria bagian dalam dari sisi matriks ke sisi sitoplasma, dan keenam proton ini akan menghasilkan tiga molekul ATP melalui penggunaan ATPase mitokondria.



