Inti: Morfologi, Struktur, Komposisi Kimia, Fungsi dan Makna Inti!
Fitur sel yang paling menonjol jika dilihat di bawah mikroskop adalah nukleus. Awalnya terdeteksi oleh Leeuwenhoek pada tahun 1700 sebagai benda yang dapat ditarik di tengah sel darah dari darah Salmon.
Struktur ini, yang pasti merupakan inti, terlihat dengan lensa sederhana yang ditemukannya di waktu luangnya. Fontana pada tahun 1781 mengamati tubuh oval serupa di dalam sel kulit belut, dan deskripsi lengkapnya diberikan oleh ahli botani Skotlandia Robert Brown pada tahun 1835.
Ini adalah struktur sel yang hampir universal pada suatu saat selama siklus hidupnya, meskipun sel-sel seperti tabung saringan tanaman vaskular dan sel darah merah mamalia dapat kehilangan nukleusnya pada saat mereka sepenuhnya berdiferensiasi.
Nukleusnya tidak homogen, tetapi mengandung struktur seperti benang yang ditemukan beberapa tahun kemudian. Struktur ini dapat diperoleh dengan pewarna anilin yang karenanya dikenal sebagai kromosom (Gr-chroma = warna dan Soma = tubuh). Sutton pada tahun 1903, pertama kali mengemukakan bahwa faktor keturunan atau gen dibawa dalam kromosom.
Dan pada tahun 1871 untuk pertama kalinya diisolasi oleh Miescher, analisis morfologisnya jauh lebih maju daripada analisis sitoplasma. Kossel mampu mendemonstrasikan keberadaan asam nukleat, dan untuk itu dia dianugerahi Hadiah Nobel pada tahun 1910.
Sel tanpa inti memiliki masa depan yang sangat terbatas. Satu-satunya jenis sel hewan yang umum tanpa nukleus, sel darah merah mamalia, hanya hidup selama 3 sampai 4 bulan; selain perannya dalam transportasi oksigen, aktivitas metabolismenya sangat terbatas.
Sel telur yang nukleusnya telah dihilangkan secara eksperimental dapat membelah untuk sementara waktu, tetapi hasil pembelahan tidak pernah berdiferensiasi menjadi jenis sel khusus, dan akhirnya mati. Fragmen tanpa nukleus, dipotong dari organisme aselular besar seperti amuba atau alga, Acetabularia, bertahan sementara, tetapi akhirnya mati kecuali nuklei dari sel lain ditransplantasikan ke dalamnya.
Nukleus sangat penting untuk kelanjutan metabolisme jangka panjang dan kemampuan sel untuk mengubah struktur dan fungsinya secara signifikan (seperti dalam diferensiasi). Sebagian besar, ini mencerminkan peran utama nukleus dalam memproduksi RNA yang dibutuhkan untuk sintesis protein.
Ketika sel berubah, fungsi dan struktur baru membutuhkan protein baru. Bahkan sel-sel yang metabolisme dan strukturnya konstan menunjukkan penggantian (perputaran) makromolekul yang terus-menerus dan mungkin organel, termasuk bagian dari mesin sintesis protein sitoplasma.
Morfologi:
Membentuk:
Secara umum nukleus cenderung berbentuk bulat, tetapi dapat berbentuk fusiform, ellipsoidal, pipih, bergantung pada bentuk dan fungsi sel. Pada sel-sel muda lebih sering berbentuk bulat dan terletak di tengah, tetapi pada sel-sel yang berdiferensiasi mungkin tergeser dan bentuknya tidak beraturan.
Nukleus dapat memanjang seperti pada epitel kolumnar dan sel otot. Dalam kasus tertentu menjadi tidak teratur dan sering diamati dalam kasus tertentu mengalami perubahan bentuk amoeboid yang lambat dalam sel hidup, misalnya sel tulang rawan, leukosit atau sel telur hewan.
Inti (Nucleus-singular) bentuk tidak teratur atau amoeboid sering terjadi pada sel yang dicirikan oleh metabolisme yang sangat aktif, dalam hal ini inti sering tidak hanya berukuran besar, tetapi menunjukkan peningkatan permukaan yang ditandai dengan pembentukan lobus, kantung, atau bahkan dalam kasus ekstrim cabang kompleks bercabang melalui sel.
Contoh ekstrim dari hal ini ditawarkan oleh kelenjar pemintalan larva serangga tertentu (Lepidoptera dan Trichoptera), di mana nukleus, awalnya berbentuk bulat, akhirnya terlihat seperti labirin dengan belitan yang menempati area yang luas di dalam sel.

Pada jenis sel tertentu, permukaan nukleus juga dapat ditingkatkan dengan memecahnya menjadi vesikel atau kariomerit yang kurang lebih terpisah, sehingga membentuk nukleus “Polimorfik” atau sarang nukleus. Inti berbentuk vesikular atau bulat umumnya terjadi pada sel jaringan sebagian besar hewan dan tumbuhan multisel. Inti masif biasanya terjadi pada sel kuman jantan hewan pada umumnya dan banyak tumbuhan rendah (pada tumbuhan coenocytic seperti Vaucharia dll.).
Posisi:
Posisi inti ditentukan oleh banyak penyebab, seperti tegangan permukaan, posisi vakuola, kerapatan relatif sitoplasma di berbagai bagian sel. Dalam sel embrio hampir selalu menempati pusat geometik.
Dalam sel tanaman yang tidak divakuola, bahkan ia menempati pusat massa sitoplasma. Pada sel adiposa, pada telur yang kaya akan kuning telur, nukleus didorong ke arah perifer oleh akumulasi paraplasma. Pada sel kelenjar terletak pada posisi basal, granula menempati sitoplasma apikal. Posisi inti juga terkait dengan fungsi sel.
Ukuran:
Pada tahun 1895 Boveri menunjukkan bahwa ukuran nukleus pada larva echinodermata bergantung pada jumlah kromosom yang dikandungnya. Gates 1901, bagaimanapun, memberikan bukti yang menunjukkan bahwa aturan ini sama sekali tidak universal. Ukuran nukleus bervariasi, tetapi secara umum memiliki beberapa hubungan dengan sitoplasma. Ini dapat dinyatakan secara numerik dalam apa yang disebut indeks nukleoplasma (NP) oleh O. Hertwigi 1906).
NP=Vn/ Vc-Vn
Dimana Vn adalah volume inti dan Vc adalah volume sel.
Indeks NP ini menunjukkan hubungan antara volume sitoplasma dan inti, yaitu jika yang pertama meningkat maka yang kedua juga harus meningkat. Rasio volume inti dengan sitoplasma dalam sel disebut ‘rasio nukleoplasma’ atau ‘rasio karioplasma’.
Nomor:
Jumlah nuklei yang ada dalam setiap massa protoplasma sangat tergantung pada massa karena dalam batas tertentu rasio permukaan inti terhadap volume sitoplasma harus dipertahankan untuk tindakan yang tepat dari keseluruhan sistem.
Umumnya semua sel adalah uninucleate atau mononucleate yaitu satu nukleus dalam satu sel tetapi dalam kasus tertentu terjadi kondisi binukleat seperti pada Paramecium caudatum, disini satu nukleus lebih kecil yaitu mikronukleus sedangkan yang lebih besar disebut makro atau meganukleus. Di beberapa negara lain, kondisi poli-atau multinukleat ada seperti di Opalina, serat otot lurik, dll.
Struktur:
Inti terdiri dari empat komponen:
(1) Selubung nuklir,
(2) Getah nuklir,
(3) Nukleolus, dan
(4) Jaringan kromatin.
1. Amplop Nuklir:
Amplop nuklir baru-baru ini menikmati banyak penyelidikan. Seperti yang dilihat dengan mikroskop kontras fase standar, tampak seperti garis gelap, garis ini memisahkan isi inti dari sitoplasma. Di sebagian besar sel, diamati untuk memecah dan membentuk kembali sebagai pusat sel dan menyelesaikan pembelahan. Namun, pada beberapa alga, protozoa, dan jamur, ia tidak terurai selama pembelahan.
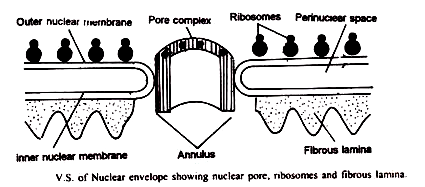
Pengamatan dengan mikroskop cahaya menunjukkan bahwa selubung nukleus berfungsi sebagai penghalang antara nukleus dan sitoplasma. Cara nukleus mengatur aliran material baru mulai dipelajari. Studi ultra-struktural mengungkapkan bahwa selubung nuklir sebenarnya terdiri dari dua membran yang dipisahkan oleh ruang perinuklear selebar 110 hingga 400 A°.
Membran bagian dalam tampaknya bersentuhan dengan kromatin inti, yang terlihat terkondensasi di sepanjang permukaannya; membran luar kontinu dengan retikulum endoplasma sitoplasma dan sering diamati dengan ribosom terikat pada permukaannya.
Hubungan membran luar dengan RE menghasilkan kontinuitas antara nukleus dan sitoplasma. Namun, kontinuitas ini tidak lengkap karena adanya membran inti dalam yang masih berfungsi sebagai penghalang struktural.
Pori-pori Nuklir :
Karena sintesis asam ribonukleat terjadi di dalam nukleus sedangkan sintesis protein terjadi di sitoplasma, jelas bahwa zat harus dapat bergerak keluar. Nyatanya, ada aliran material di kedua arah melalui pori-pori nuklir, yang, meskipun ukurannya, bukanlah saluran terbuka lebar.
Kedua membran nukleus menyatu bersama pada interval untuk membentuk pori-pori dengan diameter sekitar 600 A°. Ini pertama kali dilihat oleh Callan dan Tomlin (1950). Diperkirakan bahwa pori menempati sekitar 10% dari luas permukaan membran inti atau jumlah total pori per inti dapat bervariasi dari 100 sampai 5 x 10 7 . Pori-pori ini muncul sebagai area melingkar atau poligonal.
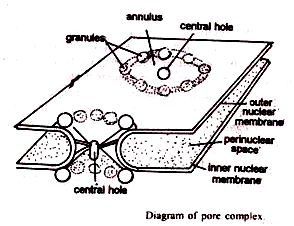
Namun, pori-pori itu bukan sekadar lubang di membran inti. Sebagian besar bukaan ditempati oleh susunan silinder atau seperti cincin (annulus) dari butiran padat sedang atau bahan fibrilar yang tampaknya bersambungan dengan lamina fibrosa bagian dalam.
Bahan annular diatur sebagai kompleks pori. Annulus terdiri dari delapan granula annular pada permukaan sisi nuklear dan sitoplasmanya dan sebuah granula pusat di tengahnya. Serat memanjang dari granul pusat dan bahan annular.
Beberapa bahan amorf membentuk diafragma di atas pori. Signifikansi diafragma dan butiran tidak pasti karena mereka tampaknya bukan konstituen universal pori-pori nuklir. Mungkin mereka adalah struktur sementara yang hadir dalam keadaan fisiologis tertentu dan tidak pada orang lain.
Lamina Berserat:
Selaput nukleus dari banyak sel menampilkan lapisan tambahan yang disebut fibrous lamina. Itu berlawanan dengan aspek dalam dari membran nuklir bagian dalam yang menghadap bagian dalam nukleus. Ini terdiri dari filamen halus yang terbuat dari protein. Mungkin mereka memberikan penguatan mekanis pada membran nuklir.
Menurut beberapa pekerja, fibrous lamina mempengaruhi pertukaran bahan antara nukleus dan sitoplasma. Tingkat perkembangan lamina berserat sangat bervariasi dalam berbagai jenis sel. Dalam sel mamalia itu tipis tetapi sangat berkembang di Amoeba dan invertebrata tertentu lainnya. Pada Amuba memiliki konfigurasi seperti sarang lebah dan memiliki ketebalan 1000-1500 A°.
Permeabilitas Amplop Nuklir :
Beberapa percobaan menunjukkan bahwa kompleks pori mungkin merupakan bukaan sementara atau permanen dalam amplop nuklir. Dengan menyuntikkan partikel emas koloid, dengan ukuran bervariasi dari 2,5 hingga 17 nm, ke dalam sitoplasma amuba, ditemukan bahwa partikel dengan diameter hingga 8,5 nm dengan cepat memasuki nukleus.
Partikel dengan diameter 8,9 hingga 10,6 nm menembus lebih lambat, dan yang lebih besar tidak masuk sama sekali. Hasil ini menunjukkan bahwa bukaan lebih kecil dari ukuran pori yang ditunjukkan. Bukti telah diperoleh dengan teknik ini, menunjukkan bahwa pori adalah jalur untuk pertukaran makromolekul. Annuli dapat mengatur pertukaran dalam kaitannya dengan ukuran dan mungkin sifat kimiawi dari zat yang menembus.
Penting untuk mempertimbangkan permeabilitas amplop nuklir tidak tetap, tetapi bervariasi dalam jenis sel yang berbeda dan dalam sel tertentu setidaknya selama siklus pembelahan. Perbedaan tersebut disebabkan perubahan sifat bahan annular (Feldher, 1971).
Kehadiran pori-pori dalam amplop nuklir harus berkorelasi dengan beberapa sifat elektrokimia dari struktur ini, yang dapat diselidiki dengan mikroelektroda garis.
Dua jenis amplop nuklir telah dikenali dengan teknik ini. Ketika sel raksasa dari kelenjar ludah Drosophila ditembus dengan mikroelektroda, terjadi perubahan mendadak pada potensial membran plasma (-12 mV); kemudian, saat mikroelektroda memasuki nukleus, ada lagi penurunan potensial negatif pada membran nukleus (-13 mV).
Hasil ini menunjukkan bahwa selubung inti mungkin merupakan penghalang difusi untuk ion sekecil K + , Na + , atau Cl. Namun, dalam amplop nuklir yang ada di oksit, tidak ada potensi yang dapat dideteksi, sehingga menunjukkan pertukaran ion bebas antara nukleus dan sitoplasma.
 Ada beberapa pengamatan morfologis yang menunjukkan adanya lintasan ribonukleoprotein dan makromolekul lainnya melintasi pori-pori inti. Bahan padat yang memanjang melalui pori-pori inti telah diamati pada oksit Amfibi. Beberapa materi mungkin sesuai dengan subunit ribosom, yang lain dengan RNA pembawa pesan.
Ada beberapa pengamatan morfologis yang menunjukkan adanya lintasan ribonukleoprotein dan makromolekul lainnya melintasi pori-pori inti. Bahan padat yang memanjang melalui pori-pori inti telah diamati pada oksit Amfibi. Beberapa materi mungkin sesuai dengan subunit ribosom, yang lain dengan RNA pembawa pesan.
Mekanisme lewatnya zat-zat ini dicapai tidak diketahui. Studi sitokimia, bagaimanapun, telah mengungkapkan adanya ATPase di pori-pori yang dapat memberikan energi yang diperlukan untuk transfer makromolekul.
Asal membran inti :
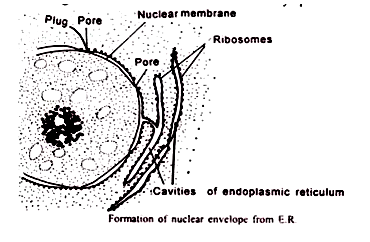
Membran nuklir dianggap sebagai struktur sitoplasma khusus yang berasal dari sistem membran sitoplasma. Selama telofase, itu terbentuk oleh akumulasi vesikel retikulum endoplasma di sekitar kelompok kromosom, yang kemudian bergabung membentuk membran lengkap.
Pada dasarnya ada dua teori tentang bagaimana selubung nukleus menyusun kembali setelah telofase, baik pasangan lamella disintesis de novo di tempat di mana kromosom dan sitoplasma bersentuhan satu sama lain (Jones 1960) atau bagian dari retikulum endoplasma yang sudah ada berdampingan. inti telofase dan bergabung untuk membentuk amplop nuklir.
Pada spermatosit belalang, misalnya, vesikel 0,5 ц yang terbentuk pada permukaan kromosom, tidak dapat dibedakan dari vesikel retikulum endoplasma (Bahr, 1959, 61). Mereka secara bertahap mengatur diri mereka sendiri sehingga sejajar dengan permukaan inti dan kemudian bergabung untuk membentuk selubung inti yang terus menerus.
2. Getah Nuklir:
Massa asidofilik yang tidak ternoda atau ringan, “karyolymph” atau “getah nuklir” mengisi sepenuhnya ruang nuklir tempat komponen lain ditemukan. Claude-lah yang pada tahun 1943 mengendapkan getah nuklir dengan fiksatif histologis dan diwarnai dengan pewarna asam. Dalam telur dan sel yang lebih besar (misalnya, Acetabularia), getah nuklir terlihat jelas. Menurut Stick (1951, 58) memberikan uji sitokimia positif untuk RNA, protein termasuk gugus-SH dan glikoprotein.
Banyak jalur metabolisme juga telah ditunjukkan dalam getah nuklir, mirip dengan sitoplasma. Ini termasuk glikolisis, shunt heksosa monofosfat, siklus asam sitrat, dll. Shunt heksosa monofosfat penting karena memasok nukleus dengan pentosa. Selanjutnya, di dalam nukleus NAD + disintesis yang merupakan koenzim dari banyak dehidrogenase. Enzim karakteristik nukleus adalah DNA dan RNA-polimerase.
Komposisi Kimia Nukleus:
Komponen kimia berikut telah dianalisis dari bahan nuklir yang diisolasi:
(i) Nukleoprotein
(a) Nukleo-protamin
(b) Nukleo-histon
(c) Non-histones atau Protein asam.
(ii) Asam nukleat.
(iii) Enzim.
(iv) Lipid.
(i) Nuceoprotein:
Selain asam nukleat, konstituen penting lainnya dari inti sel adalah komponen protein. Kromosom sel tumbuhan dan hewan tidak seperti materi genetik bakteri, terdiri dari DNA yang terkait dengan protein. Dalam protein kromosom poligen raksasa telah ditemukan di daerah band dan integrand aktivitas gen.
Protein dikaitkan dengan loop panjang kromosom sikat lampu. Nukleolus mengandung protein dalam proporsi yang tinggi dan tentu saja terdapat protein dalam getah nukleus atau nukleoplasma, dan, terkait dengan lipid, dalam membran nukleus.
Bagian protein dari nukleus tentunya sangat kompleks dan memiliki beberapa komponen. Dari jumlah tersebut yang paling dikenal adalah dua protein yang sangat mendasar dan sederhana: protamine dan sejarah. Selain itu ada protein asam, yang disebut protein dan enzim non-histone juga ada.
(a) Protamine atau Nucleo-protamine:
Miescher-lah yang menemukan protamine dalam sperma Salmon. Ini adalah protein dasar sederhana yang memiliki berat molekul sangat rendah. Ini sangat kaya akan asam amino dasar arginin dan ditemukan di spermatozoa beberapa ikan dan terikat erat dengan DNA melalui ikatan garam.
Karena protamine lebih kecil dari histones, yang berarti kromosom dan nuklei dapat dikemas ke dalam ruang yang lebih kecil, memungkinkan mobilitas sperma yang lebih besar. Secara umum, mereka terdiri dari sekitar 28 polipeptida arginin, dengan panjang total 100 A° dan mengandung 19 arginin dan 8 atau 9 asam amino nonbasa. Selama perkembangan terjadi penggantian histon secara progresif dengan protamine. Ini mungkin karena afinitas protamine yang lebih tinggi untuk DNA.
(b) Nukleo-hoston :
Histon ditemukan di eritrosit angsa oleh Kossel dan segera setelah itu ditemukan di kelenjar timus oleh Lilientfeld. Histon juga telah ditemukan pada inti sel tumbuhan tertentu, terutama embrio gandum dan kacang cedar oleh Mirsky.
Nukleoprotein juga merupakan protein dasar yang memiliki berat molekul sekitar 12000. Selain arginin sekitar 13%, histon mengandung residu dasar lainnya termasuk lisin dan histidin. Beberapa histon dengan komposisi berbeda telah diisolasi, dan tiga jenis telah dikarakterisasi.
(i) Sangat kaya lisin.
(ii) Kaya arginin.
(iii) Dan sedikit kaya lisin.
Ini heterogen dan terdiri dari beberapa komponen. Histon ditemukan di semua inti organisme tingkat tinggi, meskipun hanya sedikit dipelajari pada tumbuhan. Namun, yang ditemukan pada tanaman menyerupai histon yang ditemukan pada vertebrata.
Fungsi utama histon dianggap berasal dari bertindak sebagai “perekat” kromosom yang mengikat unit genetik DNA. Juga diketahui bahwa DNA saja, dan bahwa protein (terutama histon) dalam kompleks nukleo-protein sebagian dapat melindungi DNA dari kerusakan radiasi.
Bukti sekarang menunjukkan, bagaimanapun, bahwa peran yang sangat penting yang dimainkan oleh histon terletak pada mewakili aktivitas genetik sel. Disarankan oleh Stedman dan Stednian pada tahun 1950 bahwa histon berinteraksi dengan DNA dengan cara tertentu, mencegahnya bertindak sebagai tamplate untuk sintesis RNA dan dengan demikian mencegah mencegah transfer informasi genetik ke sitoplasma.
(c) Non-histon atau Protein Asam :
Analisis kimia nuceli terisolasi dan benang kromatin menyebabkan demonstrasi jenis protein lain yang umumnya disebut protein non-histone. Protein ini mengandung tryptophane dan memiliki sifat asam.
Fraksi non-histone lainnya telah diidentifikasi dari benang kromatin yang diisolasi. Residu yang tidak larut muncul di bawah mikroskop sebagai benang spiral yang mempertahankan karakteristik kromosom interfase. Dari sudut pandang kimiawi, sangat menarik bahwa sisa kromosom ini mengandung lebih banyak asam ribonukleat (RNA) daripada DNA.
Sejumlah besar protein non-histone hadir dalam sel yang aktif secara metabolik sangat kontras dengan komposisi spermatozoon, yang jauh lebih tidak aktif.
(ii) Asam nukleat:
Asam nukleat, yang terdiri dari dua jenis, yaitu asam ribonukleat (RNA) dan asam deoksiribonukleat (DNA) telah diamati di wilayah inti sel.
(iii) Enzim Nuklir:
Sejumlah enzim telah ditemukan dalam germinal vesikel oosit katak yang diisolasi oleh Brachet dan Dupsiva. Konsentrasi mereka yang sebenarnya sulit untuk dipastikan karena permeabilitas membran inti yang tinggi dan enzim juga cenderung bocor keluar dari vesikel germinal yang terisolasi selama isolasi mereka.
Menurut penelitian terbaru, enzim nuklir terbagi dalam dua kelas. Beberapa memiliki distribusi umum dimana yang lain ditemukan secara istimewa di jaringan tertentu. Pada kelompok pertama, hanya beberapa enzim yang berhubungan dengan metabolisme nukleosida, seperti adenosine diaminase, nucleoside phosphorylase dan guanase yang ditemukan dalam konsentrasi tinggi.
Enzim lain sebagai esterase hadir dalam berbagai konsentrasi. Yang lainnya lagi, seperti alkaline phosphatase, nucleotide phosphatases dan β-glucoronidase ada dalam konsentrasi rendah atau tidak ada sama sekali. Enzim khusus, katalase dan arginase tampaknya terkonsentrasi di beberapa nuklei tetapi kurang di nukleus lainnya.
(iv) Lipid nuklir:
Kandungan lipid nukleus telah diselidiki dalam nukleus yang terisolasi. Baru-baru ini telah dilaporkan bahwa kompleks lipo-protein dalam nuklei dari limpa sapi dan eritrosit ayam adalah sekitar 10% lipid, ini memberikan tes positif untuk fosfolipid dan kolesterol.
3. Nukleolus:
Nukleus mengandung butiran padat besar, bulat dan asidofilik yang dikenal sebagai nukleolus. Nukleolus ditemukan oleh Fontona pada tahun 1781. Pada abad ke-19 diketahui bahwa ukuran nukleolus berhubungan dengan aktivitas sintetik sel.
Oleh karena itu, sel dengan sedikit atau tanpa aktivitas sintetik, misalnya sel sperma, blastomer, sel otot, dll., ditemukan mengandung nukleolus yang lebih kecil atau tidak sama sekali, sedangkan oosit, neuron, dan sel sekretorik yang menyintesis protein atau zat lain mengandung secara komparatif. nukleolus berukuran besar.
Jumlah nukleolus dalam nukleus tergantung pada spesies dan jumlah kromosom. Jumlah nukleolus dalam nukleus dalam sel mungkin satu, dua atau empat. Posisi nukleolus dalam nukleus eksentrik.
Struktur Halus Nukleolus :
Mengenai struktur halus nukleolus, telah dilaporkan oleh Estable dan Sotelo (1955) bahwa mereka terbentuk dari dua bagian – nukleolonema berserabut dan amorf pars. Nukleolonema dikatakan membelah dan didistribusikan secara merata ke dalam mitosis anak perempuan dan dianggap sebagai struktur permanen yang bertahan selama mitosis dalam hubungannya dengan kromosom.
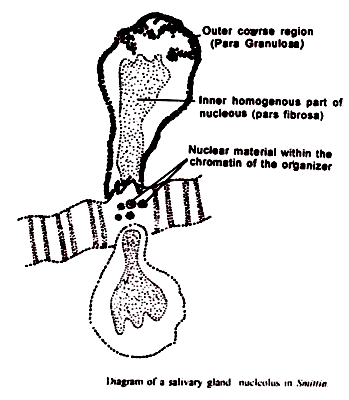
Dipercayai bahwa bagian filamen nukleolus mengandung DNA sedangkan pars amorpha sebagian terdiri dari RNA. Pars amorpha mengalami siklus karakteristik pembentukan pada telofase dan menghilang pada profase.
Pengenalan media dan teknik penyisipan baru memungkinkan analisis organisasi nukleolus yang lebih baik.
Dengan demikian empat komponen prinsip dapat dikenali:
- Bagian fibrilar dengan diameter sekitar 50A° dan panjang hingga 300-400 A° (Marrinozzi, 1963). Dalam beberapa kasus fibril ini dapat dilihat memiliki struktur beruntai ganda (Terzakis, 1965). Struktur ini juga digambarkan sebagai elemen tubular dengan diameter 15 A 0 .
- Bagian granular terdiri dari butiran padat dengan diameter rata-rata 150-200 A°, kurang lebih jumlahnya banyak di sepanjang dan di antara untaian jaringan serat (Marrinozzi, 1963).
- Daerah amorf dengan kerapatan elektron rendah, ditemukan di beberapa nukleolus (Terzakis, 1965) yang terbuat dari protein.
- Kromatin terkait nukleolus yang terletak di sekitar nukleolus dan seringkali memiliki komponen intra-nukleolus tampaknya konstan dalam sel vertebrata, tetapi jumlah atau jumlahnya masing-masing dapat bervariasi.
Kimia :
Studi sitokimia menunjukkan bahwa 5 sampai 10 persen nukleolus adalah RNA; sisanya adalah protein. Komponen protein utama adalah fosfoprotein. Tidak ada nadanya yang ditemukan di nukleolus yang terisolasi.
Ada bukti yang menunjukkan adanya ortofosfat, yang dapat berfungsi sebagai prekursor fosfor RNA. Sedikit yang diketahui tentang isi enzim nukleolus. Namun, keberadaan enzim fosfatase asam, fosforilase nukleosida dan NAD + telah ditunjukkan. RNA metilase juga telah terlokalisasi dalam nukleolus sel tertentu.
DNA tidak ada. Nukleolus dapat dikelilingi oleh cincin kromatin Feulgen-positif yang sebenarnya mewakili daerah heterokromatik kromosom.
Biogenesis Nukleolus atau Siklus Nukleolus:
Nukleolus sebagai badan yang terorganisir tidak memiliki kontinuitas. Ini menghilang pada awal pembelahan sel (profase) dan muncul kembali pada akhir pembelahan sel pada tahap telofase.
Nukleolus terbentuk pada daerah tertentu oleh satu atau lebih kromosom dari satu set kromosom haploid. Kromosom semacam itu dikenal sebagai kromosom nukleolar.
Mayoritas spesies diploid memiliki dua kromosom nukleolar di setiap sel diploid atau somatik. Namun pada manusia kromosom berjumlah 13, 14, 15, 21 dan 22 ikut serta dalam pembentukan nukleolus.
Wilayah spesifik dari kromosom ini yang aktif dalam pembentukan nukleolar dikenal sebagai zona nukleolar organisator nukleolar. Sangat sering tetapi tidak selalu, ditandai dengan penyempitan sekunder. Pengatur nukleolar membawa gen untuk RNA ribosom 18S dan 28S.
Penyempitan sekunder dengan pengatur nukleolar secara morfologis berbeda dengan penyempitan sekunder lainnya. Pada tahap pachytene dari Zea mays, badan pengorganisasian nukleolar bernoda gelap dikaitkan dengan zona nukleolar.
Jenis nukleolus :
Atas dasar distribusi butiran dan fibril ribonukleoprotein, tiga jenis nukleolus dapat dibedakan (Wilson).
- Nukloli dengan nukleolonema, yang terdapat pada sebagian besar sel yang disebut plasmosom.
- Nukleolus kompak tanpa nukleolonema, yang telah dijelaskan di kelenjar ludah sciurid dan di protozoa, Tetrahymena pyriformis. Dalam nukleolus ini butiran dan fibril ribonukleoprotein terdistribusi secara merata.
- Nukleolus berbentuk cincin dengan butiran ribouncleoprotein dan fibril hanya ada di daerah perifer (nudeolonema perifer) telah dilaporkan dalam sel endotel, sel otot polos dan sel sarkoma limfo. Dalam nukleolus ini, wilayah tengah sebagian besar terdiri dari kromatin.
Fungsi:
(i) produksi RNA:
Nukleolus adalah salah satu situs sintesis RNA yang paling aktif. Ini menghasilkan sekitar 70-90 persen RNA seluler di banyak sel. Ini adalah sumber RNA ribosom (rRNA). Kromatin dalam nukleolus mengandung gen atau DNA ribosom (rDNA) untuk mengkode RNA ribosom.
Fibril mewakili asal RNA ribosom dan butiran tahap selanjutnya. Butiran pada gilirannya adalah prekursor ribosom. Nukleolus sehingga membuat prekursor ribosom, bukan seluruh ribosom.
Kromasi → Fibril → Butiran → Ribosom
(Mengandung DNA) (Mengandung RNA) (Mengandung RNA).
Nukleolus juga dapat menghasilkan beberapa jenis messenger RNA (mRNA), dan setidaknya satu jenis RNA dengan berat molekul rendah.
(ii) Sintesis protein:
Maggio (1960) dan lainnya telah menyarankan bahwa sintesis protein terjadi di nukleolus. Jika ini benar maka protein ribosom diproduksi di nukleolus. Studi lain, bagaimanapun, menunjukkan bahwa protein ribosom disintesis dalam sitoplasma.
(iii) Formasi Ribosom:
Pada eukariota, pengkodean gen untuk RNA mengandung rantai setidaknya 100 hingga 1.000 salinan DNA yang berulang. DNA ini dilepaskan dalam bentuk loop dari serat kromosom. Loop DNA dikaitkan dengan protein untuk membentuk nukleolus.
DNA berfungsi sebagai cetakan untuk 45S rRNA. Setengah dari 45S rRNA dipecah menjadi 28S dan 18S rRNA. Setengah lainnya dipecah lebih lanjut ke tingkat nukleotida. Di dalam nukleolus, 28S rRNA bergabung dengan protein yang dibuat dalam sitoplasma untuk membentuk subunit ribosom 60S. 18S rRNA juga berasosiasi dengan protein untuk membentuk subunit 40S ribosom.
- Jaringan Kromatin :
Kromatin muncul sebagai struktur seperti benang, melingkar dan memanjang. Ini diwarnai dengan pewarna dasar seperti pewarna Feulgen, acetocarmine dll., Oleh karena itu disebut sebagai serat kromatin atau zat kromatin (Gr., chromecolour, soma=tubuh).
Ini terlihat selama tahap interfase. Selama pembelahan sel mereka menjadi struktur seperti pita tebal yang dikenal sebagai kromosom.
Signifikansi Nukleus:
Eksperimen Hammerling:
- Hammerling, seorang ahli biologi Jerman, secara eksperimental membuktikan pentingnya nukleus. Jika sebuah fragmen yang mengandung nukleus dipotong dari Acetabularia dari satu spesies, yang dicirikan oleh morfologi tertentu, fragmen tersebut akan meregenerasi seluruh sel dari spesies tersebut.
Kemampuan regeneratif ini memungkinkan eksperimen dari jenis yang diilustrasikan pada Gambar 8.6 di mana inti dari satu spesies digabungkan dengan sitoplasma dari spesies yang berbeda. Hammerling melakukan eksperimen tertentu dengan menggunakan dua spesies alga hijau, Acetabularia. Dua spesies yaitu A. crenulated dan A, Mediterranean yang digunakan dalam percobaan ini berbeda pada bentuk tudungnya. Sementara pada A. crenulata tutupnya memiliki sinar lepas, pada A. mediterranea ditemukan tutup seperti payung.
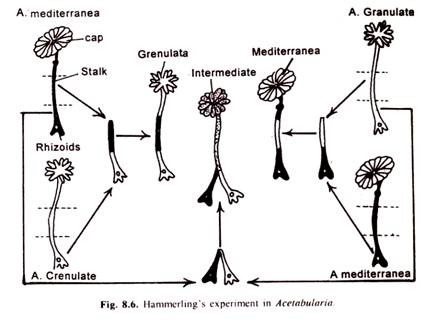
Nukleus pada kedua spesies terletak di rizoid di bagian bawah tangkai. Jika tutupnya dipotong, maka akan berkembang lagi dan bentuknya akan seperti tipe aslinya. Namun, jika setelah melepas tudung, tangkai satu spesies dicangkokkan pada rizoid (yang mengandung nukleus) spesies lain, bentuk tudung akan ditentukan oleh nukleus dan bukan oleh tangkai. Jika nukleus milik A. crenulata, bentuk tudung akan menjadi tipe crenulata. Jika nukleus berasal dari A. mediterranea, tudung akan bertipe mediterranea.
Ketika kedua inti hadir, bentuk tudung akan menjadi perantara. Kesimpulan yang ditarik dari percobaan tersebut adalah bahwa nukleus menghasilkan bahan yang masuk ke sitoplasma dan berpartisipasi dalam pengendalian pertumbuhan sel dan morfologi sel.
Temuan penting adalah bahwa morfologi sel yang beregenerasi hampir menjadi seperti spesies dari mana nukleus diambil. Pada fragmen hibrid dengan nukleus dari satu spesies dan sebagian besar sitoplasma dari spesies lain, bahan sitoplasma lama bertahan untuk sementara dan dapat mempengaruhi bentuk sel. Namun, akhirnya, ini habis dan digantikan oleh bahan yang baru diproduksi dari nukleus.



