Catatan singkat tentang Laki-laki Heterogamet dan Betina Heterogamet – Biologi Sel!
(A) Laki-laki heterogami:
Jenis kelamin perempuan memiliki 2 kromosom ‘X’ (XX) sedangkan laki-laki hanya memiliki satu kromosom ‘X’ dan pada saat gametogenesis menghasilkan 2 jenis gamet. 50% dengan kromosom ‘X’ sedangkan 50% lainnya tanpa kromosom ‘X’.
Gambar Courtesy: friendshipcircle.org/blog/wp-content/uploads/2012/05/Chromosomes.jpg
Karena 2 jenis gamet yang dihasilkan oleh jantan maka disebut kelamin heterogami (jantan). Sedangkan betina bersifat homogami.
Laki-laki heterogami terdiri dari dua jenis:
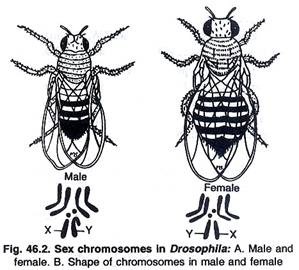
(i) Tipe XX-XY:
Pada manusia, mamalia lain, tumbuhan dan banyak serangga seperti Drosophila, dll., betina memiliki tipe XX homogami sedangkan jantan memiliki kromosom X dan Y. Betina hanya menghasilkan satu jenis gamet sedangkan jantan menghasilkan dua jenis gamet ‘X’ dan ‘Y’. Jenis kelamin embrio tergantung pada jenis sperma atau gamet jantan (tipe X atau Y). Jika gamet betina dibuahi oleh sperma ‘X’ maka embrio akan berjenis kelamin betina sedangkan jika dibuahi oleh sperma ‘Y’ maka embrio akan menjadi jantan (Gambar 46.2).
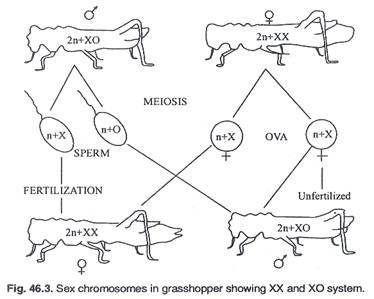
(ii) Tipe XX-XO:
Pada beberapa serangga seperti belalang, serangga, dll., betina adalah tipe XX homogami tetapi jantan hanya memiliki satu kromosom seks, yaitu ‘X’ saja, tidak ada kromosom ‘Y’; maka jumlah kromosom pria dan wanita akan berbeda. Laki-laki memiliki kromosom nomor satu lebih sedikit daripada perempuan. Betina akan menghasilkan hanya satu jenis telur (‘X’) tetapi jantan menghasilkan 50% sperma dengan ‘X’ dan 50% lainnya tanpa ‘X’. Jenis kelamin anak tergantung pada sperma yang membuahi sel telur (Gambar 46.3).
(B) Betina heterogami:
Pada sebagian besar spesies seperti burung, ngengat, dan beberapa ikan, terjadi mekanisme penentuan jenis kelamin kromosom yang pada dasarnya identik dengan mekanisme XX-XY. Karena betina heterogami, maka ditetapkan sebagai ZW. Laki-laki homogami, mereka ditetapkan sebagai ZZ. Ini disebut sebagai tipe ZZ-ZW, bukan sistem XX-XY.
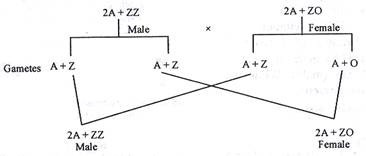
(i) tipe ZO-ZZ:
Betina memiliki kromosom Z tunggal pada ngengat, kupu-kupu, dan ayam peliharaan. Betina menghasilkan dua jenis telur satu dengan kromosom ‘Z’ lainnya tanpa ‘Z’. Namun, pejantan hanya menghasilkan satu jenis sperma yang membawa ‘Z’ saja. Jenis kelamin ditentukan berdasarkan jenis sel telur yang dibuahi oleh sperma jika mengandung Z maka embrio akan jantan jika tidak maka embrio akan betina.
(ii) tipe ZW-ZZ:
Ini umum terjadi pada serangga, vertebrata seperti reptil ikan, burung, dll. Betina heterogami dengan ZW dan jantan homogami dengan ZZ. Betina menghasilkan dua jenis sel telur, yaitu 50% dengan ‘Z’ atau 50% dengan ‘W’ sedangkan jantan hanya menghasilkan satu jenis sperma, yaitu dengan ‘Z’. Jenis kelamin keturunan tergantung pada jenis sel telur yang dibuahi.
Kromosom ‘Y’ dalam Penentuan Jenis Kelamin:
Pada Drosophila dan pria, wanita normal memiliki kromosom XX dan pria memiliki kromosom XY. Kromosom ‘X’ mengandung gen keperempuanan tetapi kromosom ‘Y’ mengandung gen kelelakian. Pada mamalia, keberadaan kromosom ‘Y’ diperlukan untuk perkembangan fenotip seks laki-laki. Pada Drosophila, kromosom ‘Y’ tidak berperan dalam penentuan jenis kelamin.
Jumlah kromosom seks yang tidak beraturan cukup umum terjadi pada manusia. Jika kromosom seks ‘X’ hadir dalam nomor apa pun, misalnya, XXX atau XXXX, dll., dengan tidak adanya kromosom ‘Y’ memunculkan fenotipe seks wanita. Kromosom ‘Y’ diperlukan untuk kejantanan, misalnya XXXXY. Kromosom ‘Y’ menginduksi perkembangan medula gonad yang tidak berdiferensiasi menjadi testis, sedangkan komponen kromosom XX menginduksi korteks gonad yang tidak berdiferensiasi untuk mengembangkan ovarium.
Gen pada kromosom ‘Y’ pada manusia yang bertanggung jawab untuk perkembangan testis disebut ‘TDF’ (faktor penentu testis). Dengan tidak adanya gen TDF, fenotipe seks wanita akan diekspresikan. Bahkan dengan adanya tiga atau lebih kromosom ‘X’, satu kromosom ‘Y’ biasanya cukup untuk menghasilkan testis dan karakteristik laki-laki.
Dalam album Melandrium (tipe XY) Warmke dan Westergaard dan lainnya telah menunjukkan bahwa jenis kelamin ditentukan oleh keseimbangan antara gen penentu laki-laki pada kromosom ‘Y’ dan gen penentu perempuan pada ‘X’ dan autosom. Pada tumbuhan ini, yaitu individu Dioecious ‘XY’ adalah staminate dan tumbuhan ‘XX’ adalah pistillate. Kromosom ‘Y’ adalah yang terbesar dan paling mencolok (Gbr. 46.IB). Tiga wilayah berbeda dari pengaruh kromosom ini dalam penentuan jenis kelamin dan kesuburan pria.
Wilayah I menekan keperempuanan, jika tidak ada tumbuhan bersifat biseksual, yaitu mengekspresikan kejantanan dan keperempuanan.
Wilayah II mempromosikan perkembangan jantan ketika wilayah ini hilang dengan atau tanpa wilayah I, tanaman betina diproduksi.
Wilayah III membawa gen kesuburan laki-laki hilangnya wilayah ini mengakibatkan mandul laki-laki. Hanya sebagian dari kromosom ‘Y’ (Wilayah IV) yang homolog dengan ‘X’ tetapi sebagian besar dari ‘X’ dibedakan tanpa pasangan struktural pada ‘Y Westergaard menemukan bahwa autosom juga terlibat dalam penentuan wanita.
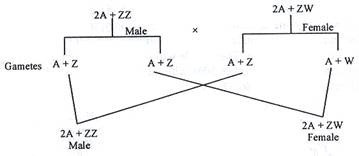
Konsep Genie Balance Penentuan Jenis Kelamin Dalam Drosophila:
CB Bridges menunjukkan bahwa penentu perempuan terletak pada kromosom ‘X’ dan penentu laki-laki berada di autosom. Tidak ada lokus spesifik yang diidentifikasi pada saat itu. Tetapi bukti terbaru menunjukkan bahwa banyak segmen kromosom yang terlibat secara khusus, gen penentu wanita ditunjukkan dibawa pada kromosom ‘X’ dan gen penentu pria ditunjukkan terletak pada tiga kromosom autosom Drosophila. Teori keseimbangan gen penentuan jenis kelamin dirancang untuk menjelaskan mekanisme penentuan jenis kelamin pada D. melanogaster. (Gbr. 46.4)
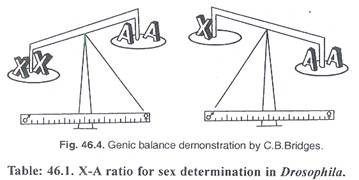
Tabel: 46.1. Rasio XA untuk penentuan jenis kelamin pada Drosophila:
|
kromosom ‘X’ |
kromosom ‘A’ |
Rasio X/A |
Seks |
|
1X |
2A |
0,5 |
Pria |
|
2X |
2A |
1.0 |
Perempuan |
|
3X |
2A |
1.5 |
Metafemale |
|
4X |
3A |
1.33 |
Metafemale |
|
4X |
4A |
1.0 |
Betina tetraploid |
|
3X |
3A |
1.0 |
Wanita triploid |
|
3X |
4A |
0,75 |
Interseks |
|
2X |
3A |
0,67 |
Interseks |
|
2X |
4A |
0,5 |
Laki-laki tetraploid |
|
IX |
3A |
0,33 |
Meta laki-laki |
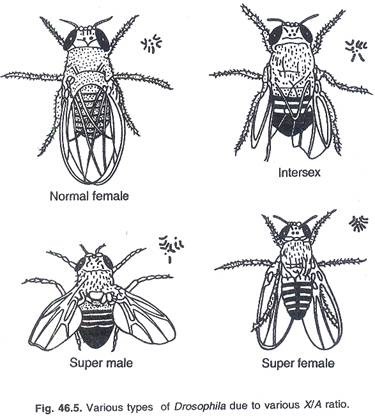
Bridges secara eksperimental menghasilkan berbagai kombinasi kromosom X dan autosom (A) di Drosophila dan menyimpulkan dari perbandingan bahwa satu kromosom ‘X’ dan dua set autosom (A) menghasilkan laki-laki normal. Laki-laki normal memiliki rasio kromosom ‘X’ terhadap kumpulan autosom sebesar 0,5 (Tabel 46.1) (Gambar 46.5).
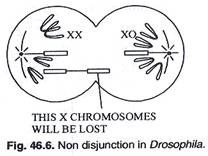
Susunan kromosom tidak beraturan pertama dari percobaan Bridges dihasilkan dari non disjungsi, yaitu kegagalan pasangan kromosom untuk berpisah secara anafase. Kromosom X, yang biasanya berpasangan selama profase meiosis oogenesis dan terpisah ke kutub yang berlawanan selama anafase, tetap bersama dan bermigrasi ke kutub yang sama.
Akibatnya beberapa gamet betina menerima 2 kromosom X dan yang lain tidak menerima kromosom X (Gambar 46.6). Setelah pembuahan oleh sperma dari pejantan tipe liar (2A + XY), semua zigot memiliki 2 set autosom (2A) tetapi beberapa menerima 2X (XX) dari ibu dan satu X dari ayah dan menjadi 3X (XXX). Rasio kromosom A/X adalah 3:2 dan lalatnya metafemale. Laki-laki XO steril dan laki-laki dengan kromosom aY dan tanpa kromosom X tidak dapat bertahan hidup (Gambar 46.7).
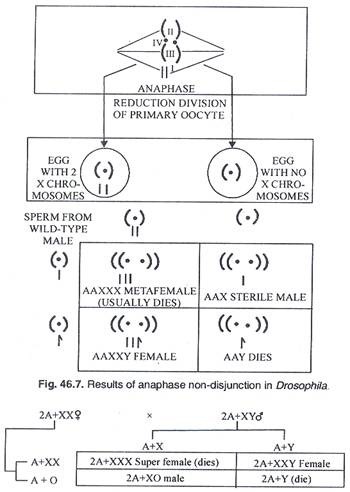
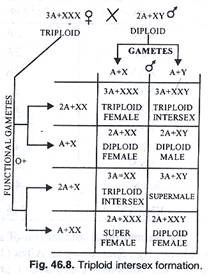
Kehadiran interseks triploid dalam percobaan yang dilakukan oleh Bridges (Gambar 46.8) merupakan bukti nyata adanya faktor pembawa autosom dalam penentuan jenis kelamin. Menurutnya (teori keseimbangan gen) rasio jumlah kromosom ‘X’ dan jumlah set lengkap autosom menentukan jenis kelamin.
Penentu perempuan terletak pada kromosom X dan penentu laki-laki pada autosom. Bukti terbaru menunjukkan bahwa banyak segmen kromosom yang terlibat dalam proses ini. Gen penentu betina dibawa pada ‘X’ dan gen penentu jantan terletak di tiga autosom Drosophila.
Haploidiploidi dan penentuan jenis kelamin pada Hymenoptera:
Anggota Hymenoptera antara lain semut, lebah, tawon, lalat gergaji, dll. Pada beberapa spesies, pejantan berkembang secara partenogenetik (dari telur yang tidak dibuahi) dengan jumlah kromosom haploid (16 pada lebah madu drone Apis mellifera). Lebah madu ratu dan pekerja muncul dari telur yang telah dibuahi dengan jumlah kromosom diploid (32). Karena jantan normal bersifat haploid dan betina bersifat diploid, mekanisme penentuan jenis kelamin disebut haplodiploidi (Gambar 46.9).
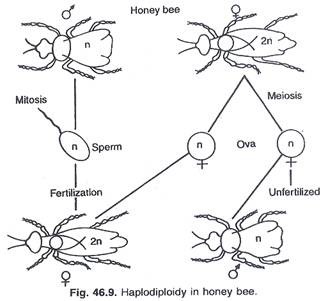
Hasil percobaan Whiting menunjukkan bahwa status homozigot, heterozigot atau hemizigot (gen dalam dosis tunggal) segmen kromosom tertentu mengontrol penentuan jenis kelamin. Penentuan betina tergantung pada heterozigositas untuk bagian dari kromosom. Jika hemizigot terbentuk, mereka akan menjadi laki-laki.
Teori Keseimbangan Genie tentang Penentuan Jenis Kelamin pada Tumbuhan:
- Westergaard adalah orang pertama yang mendemonstrasikan penentuan jenis kelamin pada tanaman dengan mempertimbangkan rasio X/A. RP Roy juga mempelajari secara detail penentuan jenis kelamin pada Coccinia dan Melandrium (tanaman berkelamin tunggal). Pada tumbuhan ini, kromosom ‘Y’ sangat penting. Hanya kehadiran kromosom ‘Y’ yang menjadikannya laki-laki. Rasio X/A tidak ada hubungannya, jika ada satu kromosom ‘Y’ saja dalam kondisi diploid atau poliploid tanaman akan menunjukkan karakter jantan.
Tabel 46.2. Penentuan jenis kelamin di Coccinia dan Melandrium:
|
kromosom X |
Kromosom |
rasio X/A |
Seks |
|
XX |
2A |
1.00 |
Perempuan |
|
XY |
2A |
0,50 |
Pria |
|
XXY |
2A |
1.00 |
Pria |
|
XYY |
2A |
0,50 |
Pria |
|
XXY |
3A |
0,67 |
Pria |
|
XXX |
3A |
1.00 |
Perempuan |
|
XY |
4A |
0,25 |
Pria |
|
XXY |
4A |
0,50 |
Pria |
|
XXXY |
4A |
0,75 |
Pria |
|
XXXXY |
4A |
1.00 |
Banci |
Mekanisme penentuan jenis kelamin kromosom seks juga telah diamati pada lumut monoploid, misalnya Spaerocarpos. Allen (1919) menemukan bahwa sporofit Spaerocarpos mengandung dua kromosom seks (XY) dan menghasilkan dua jenis meiospora (tipe X dan Y) meiospora ‘X’ berkecambah menjadi gametofit betina dan meiospora ‘Y’ berkecambah menjadi gametofit jantan.
Mosaik dan Gynandromorph:
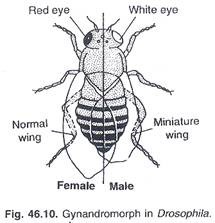
Perilaku abnormal kromosom pada serangga dapat mengakibatkan pembentukan ‘gynandromorph’ atau mozaik seksual, dimana separuh bagian hewan adalah jantan dan separuh lainnya adalah betina. Gynandromorph pada Drosophila (Gbr.46.10) adalah interseks bilateral, dengan pola warna jantan, bentuk tubuh dan jengger pada separuh tubuh dan ciri betina pada separuh lainnya. Baik gonad dan alat kelamin pria dan wanita hadir.
Ginandromorf seperti itu adalah hasil dari ketidakteraturan dalam mitosis pada pembelahan pertama zigot. Kromosom tertinggal dalam pembelahan dan tidak tiba di kutub pada waktunya untuk dimasukkan ke dalam inti yang direkonstruksi. Ketika salah satu kromosom ‘X’ dari zigot XX (betina) tertinggal di gelendong, satu inti anak hanya menerima satu kromosom ‘X’.
Pola tubuh mosaik terbentuk satu nukleus dalam tahap dua nukleus adalah XX (betina) dan yang lainnya adalah XO (jantan). Ini berkembang menjadi satu-setengah sebagai perempuan dan yang lainnya sebagai laki-laki. Pada Br aeon bebetor, gynandromorphs dapat terjadi pada bidang anterior posterior yang memunculkan tawon dengan susunan aneh seperti kepala jantan dengan perut betina atau kepala betina dengan perut jantan.
Faktor lingkungan dan penentuan jenis kelamin:
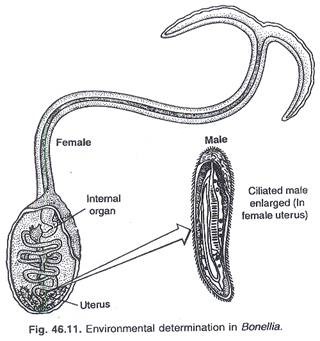
Pada beberapa hewan tingkat rendah, penentuan jenis kelamin bersifat non-genetik dan bergantung pada faktor lingkungan luar. Genotipe laki-laki dan perempuan adalah sama. Rangsangan dari lingkungan membantu memulai perkembangan ke arah satu jenis kelamin atau yang lain, misalnya cacing laut jantan Bonellia berukuran kecil dan merosot dan hidup dalam saluran reproduksi betina yang lebih besar (Gambar 46.11).
Semua organ tubuhnya merosot kecuali sistem reproduksi. F Baitzer menemukan bahwa cacing muda yang dipelihara dari satu telur terisolasi menjadi betina. Cacing yang baru menetas dalam air berisi betina dewasa menempel pada belalai betina berubah menjadi jantan, dan akhirnya bermigrasi ke saluran reproduksi betina sebagai parasit. Hal ini menunjukkan bahwa ekstrak belalai betina mempengaruhi cacing muda menjadi jantan.
Pada beberapa reptil, suhu pada saat inkubasi sebelum menetas berperan besar dalam menentukan jenis kelamin keturunan. Telur yang menetas pada suhu 26-27°C berkembang menjadi betina dan pada suhu 29°C menjadi jantan. Pada penyu (Chrysema picta) inkubasi pada suhu tinggi (lebih dari 30°C) menghasilkan betina dan pada suhu rendah menjadi jantan. Pada biawak (Agama agama) suhu inkubasi yang tinggi menghasilkan pejantan.
Meskipun pemisahan gen dan kromosom penentu jenis kelamin tertentu bertanggung jawab atas fenotip jenis kelamin pada sebagian besar spesies, potensi genetik untuk kelelakian dan keperempuanan ada di setiap zigot, namun beberapa faktor spesifik di lingkungan memicu ekspresi gen yang menghasilkan fenotipe laki-laki atau fenotipe perempuan.



