Inilah makalah penelitian Anda tentang genetika manusia, kromosom dan gen!
Kami mewarisi beberapa karakter fisik dan biokimia dari orang tua dan leluhur kami. Transmisi karakter atau sifat yang diwariskan secara turun-temurun dikenal dengan hereditas. Genetika adalah cabang biosains yang mempelajari prinsip-prinsip dasar hereditas.

Sumber gambar: mdsalaries.com/wp-content/uploads/2011/11/shutterstock_61775431.jpg
Telah ditetapkan bahwa karakter atau sifat herediter ditransmisikan oleh gen kromosom. Ekspresi karakter yang diwariskan, bagaimanapun, dimodifikasi oleh lingkungan di mana individu tumbuh dan berkembang.
Oleh karena itu, pengetahuan dasar tentang kromosom dan gen manusia sangat penting untuk memahami prinsip-prinsip genetika.
Kromosom:
Kromosom adalah struktur seperti benang yang diwarnai dalam inti setiap sel hewan. Gen ditanggung oleh kromosom dalam seri linier sebagai bagian dari molekul DNA tertentu. Kromosom individu terlihat di bawah mikroskop hanya selama pembelahan sel.
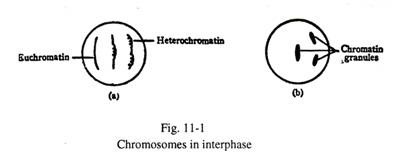
Selama interfase sel, nukleus mengandung jaringan benang atau butiran kromatin tetapi bukan kromosom individual, karena setiap kromosom menjadi terurai menjadi benang tipis panjang yang berada di luar resolusi mikroskop cahaya. Tetapi beberapa kromosom tetap melingkar di beberapa tempat dan ini diidentifikasi sebagai butiran kromatin pada interfase (Gbr. 11-1).

Bagian kromosom yang tidak digulung dikenal sebagai eukromatin yang aktif secara genetik; bagian yang digulung disebut heterokromatin yang secara genetik inert. Selama pembelahan sel, setiap kromosom melingkar rapat sepanjang panjangnya dan menjadi lebih pendek dan lebih tebal. Pada akhirnya kromosom-kromosom individu dapat dengan mudah terlihat di bawah mikroskop (Gbr. 11-2).
Oleh karena itu, kromosom secara genetik tidak aktif selama pembelahan sel. Semua aktivitas biokimia kromosom berupa replikasi DNA, pembentukan mRNA dan sintesis protein berlangsung selama interfase yang terdiri dari tiga tahap siklus sel—G 1 (Gap 1), S (Sintesis), G 2 (Gap 2 ) tahapan. Replikasi DNA terjadi pada tahap S dan mencakup periode sekitar 7 jam.
Setiap kromosom menyajikan penyempitan primer yang dikenal sebagai sentromer atau kinetocore yang melekat pada gelendong achromatic selama pembelahan sel dan mengatur pembentukan mikrotubulus kromosom (Gbr. 11-3a).

Pada profase pembelahan sel, setiap kromosom membelah secara longitudinal menjadi dua kromatid kecuali pada sentromer (Gbr. 11-3b). Ujung kromatid yang bebas dikenal sebagai telomer, yang bila utuh, tidak memungkinkan fusi dengan kromatid kromosom yang berdekatan . Kromatid dari beberapa kromosom menunjukkan konstriksi sekunder di dekat salah satu ujungnya, dan segmen kromatid di bagian distal dari konstriksi membentuk badan satelit (Gbr. 11-3c, d). Penyempitan sekunder diyakini mengatur pembentukan nukleolus.
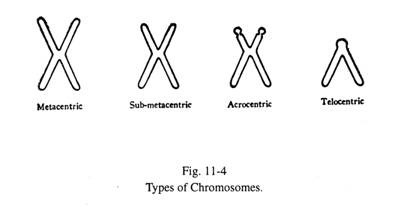
Jenis Kromosom (Gbr. 11 -4):
Sentromer menempati posisi variabel dalam kaitannya dengan pasangan kromatidnya. Dengan demikian, kromosom dapat disebut metasentrik ketika sentromer berada di tengah, sub-metasentrik ketika sentromer sedikit bergeser dari tengah, akrosentrik ketika letaknya dekat dengan ujung, dan telosentris jika sentromer menempati ujung kromosom. . Kromosom telosentris tidak ada pada manusia, kecuali secara patologis. Sebagian besar kromosom akrosentrik memperlihatkan badan satelit pada lengan pendeknya yang dipisahkan oleh penyempitan sekunder. Lengan kromatid yang lebih pendek dilambangkan dengan p dan lengan yang lebih panjang dengan q.
Jumlah Kromosom:
Jumlah kromosom konstan dalam suatu spesies. Pada manusia, jumlahnya 46 (diploid ) pada semua sel somatik dan sel germinal imatur, tetapi jumlahnya 23 (haploid) pada sel germinal atau gamet dewasa. Jumlah persis 46 kromosom pada setiap sel somatik manusia normal pertama kali terdeteksi oleh Tjio dan Levan (1956) dengan munculnya kultur jaringan.
Beberapa kelainan herediter berhubungan dengan perubahan jumlah kromosom. Ketika jumlah bertambah banyak dari kromosom haploid (23) (selain jumlah diploid), kondisi ini dikenal sebagai poliploidi. Jika poliploidi, katakanlah triploidi atau tetraploidi, memengaruhi semua sel somatik, tingkat kelangsungan hidup buruk. Poliploid pada tahap zigot dapat terjadi karena pembuahan satu ovum oleh lebih dari satu spermatozoa.
Dalam kondisi normal, poliploidi dapat ditemukan di beberapa sel hati dan di mukosa kandung kemih. Hal ini dapat terjadi pada telofase mitosis, ketika setelah terbentuknya dua membran nukleus yang menyelubungi jumlah kromosom diploid, sitoplasma gagal membelah dan kedua membran nukleus melebur yang menyelubungi kromosom diploid berjumlah ganda.
Aneuploidy adalah suatu kondisi dimana jumlah kromosom diubah oleh satu atau lebih, tetapi tidak oleh kelipatan haploid. Sebagian besar bahaya nomor kromosom terjadi pada anafase. Setelah pemisahan sentromer, satu atau lebih kromosom gagal bermigrasi dengan baik karena fungsi abnormal dari gelendong akromatik. Fenomena ini dikenal sebagai nondisjunction.
Akibatnya, kedua anggota pasangan tertentu pergi ke satu sel anak yang menerima kromosom ekstra (trisomi), dan sel anak lainnya kekurangan kromosom tersebut (monosomi). Kadang-kadang setelah pemisahan sentromer, salah satu anggota kromosom yang baru terbentuk terpisah seperti biasa membentuk komplemen kromosom normal dalam satu sel anak, sedangkan anggota lainnya gagal mencapai kutub yang berlawanan dari gelendong yang mengakibatkan kekurangan kromosom tersebut (monosomi) di bagian lain. sel putri. Ini dikenal sebagai kelambatan anafase.
Non-disjungsi dapat terjadi pada mitosis atau meiosis dan mungkin melibatkan kromosom seks serta autosom. Non-disjungsi autosomal kurang layak, terutama ketika mempengaruhi kromosom besar. Tubuh kita lebih toleran terhadap sel trisomik daripada monosomik. Sel-sel monosomik mengalami degenerasi dini. Sindrom Turner pada wanita dengan konstitusi kromosom XO 45 mungkin merupakan satu-satunya contoh individu monosomik yang hidup. Jika non-disjuncion terjadi pada pembelahan pertama zigot, maka semua sel adalah aneuploid dan individu tersebut menunjukkan mosaikisme dengan setengah dari total sel menjadi trisomik dan setengah lainnya monosomik.
Ketika non-disjunction terjadi pada meiosis I, keempat gamet menjadi abnormal (dua dengan 24 kromosom, dan dua dengan 22 kromosom ). Jika terjadi pada meiosis II, dua gamet normal dan dua abnormal. Ketika pembuahan terjadi antara gamet normal dan abnormal, semua sel organisme yang berasal dari zigot tersebut adalah aneuploid. Nondisjungsi dalam gametogenesis kadang-kadang diamati pada wanita lanjut usia (35 tahun ke atas). Mungkin oosit primer yang memulai pembelahan meiosis pertama dalam kehidupan prenatal, menyelesaikan proses sebelum ovulasi setelah selang waktu yang lama sekitar 40 tahun lebih. Penyelesaian meiosis pertama oosit yang tertunda mungkin mendukung non-disjungsi.
Susunan Kromosom:
Empat puluh enam kromosom dalam setiap sel somatik manusia normal tersusun dalam 23 pasang. Dua puluh dua pasang dikenal sebagai autosom, gen yang mengatur karakter tubuh ; pasangan yang tersisa dikenal sebagai kromosom seks yang terutama mengatur karakter seks. Satu anggota dari setiap pasangan berasal dari ayah, dan anggota lainnya berasal dari ibu.
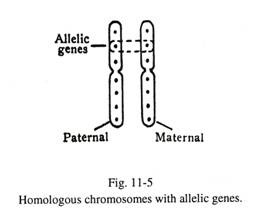
Pasangan terjadi antara kromosom identik yang identik dalam panjang, posisi sentromer, pola pita, dan distribusi gen. Kromosom berpasangan dikenal sebagai kromosom homolog (Gbr. 11-5).
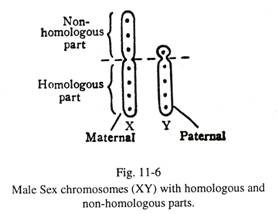
Pada wanita, kedua kromosom seks memiliki panjang yang identik dan dilambangkan dengan XX. Pada laki-laki, pasangan kromosom seks tidak sama panjangnya dan dilambangkan dengan XY. Yang lebih panjang diwakili oleh X, dan yang lebih pendek oleh Y. Selama pasangan kromosom seks laki-laki, keduanya memiliki bagian homolog dan non-ho-molog (Gbr. 11-6).
Gen atau sistron, yang merupakan bagian dari molekul DNA spesifik, terkandung di dalam kromosom dalam rangkaian linier. Mereka membentuk unit fungsional karakter turun-temurun. Posisi suatu gen dalam kromosom disebut lokusnya yang disebut dengan sentromer.
Gen tidak mengubah lokus, kecuali dalam pergantian morfologi kromosom atau dalam rekombinasi karena persilangan pada meiosis. Gen yang menempati lokus identik pada sepasang kromosom homolog dikenal sebagai alelomorf atau alel (Lihat Gambar 11-5). Gen alelik mengatur karakter fisik dan biokimia spesifik yang berbeda, melalui pembentukan RNA dan biosintesis protein.
Dalam persiapan kromosom dari kultur sel mitosis (setelah menghentikan pembelahan sel pada fase metÂ), pasangan kromosom homolog tidak divisualisasikan. Pasangan homolog hanya dicocokkan selama kariotipe dari fotomikrograf yang diperbesar. Namun, pada tahap zigoten dari profase pembelahan meiosis pertama, kromosom homolog ditemukan berpasangan yang membentuk hubungan titik ke titik; fenomena ini dikenal sebagai sinapsis.
Sex Chromatin atau badan Barr:
Selama interfase, sel somatik wanita normal memperlihatkan badan heterochromatin plano-con vex di bawah membran nukleus. Ini dikenal sebagai kromatin seks atau tubuh Barr. Ini pertama kali terdeteksi oleh Barr dan Bertram pada tahun 1949 di inti sel saraf frenikus kucing betina. Dari dua kromosom X pada wanita normal, salah satunya sangat melingkar dan anggota lainnya sangat tidak melingkar. Kromosom X inaktif secara genetik yang sangat melingkar membentuk badan Barr, yang terpampang di bawah membran inti (Gbr. 11-7).
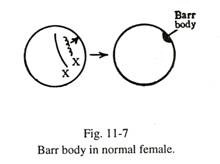
Badan-badan ini membantu dalam sexing nuklir jaringan. Tubuh Barr mudah ditemukan di sel-sel itu, yang memiliki nukleus berwajah terbuka. Biasanya badan Barr dipelajari dari sel-sel apusan bukal, atau dengan mengamati badan ‘drum stick’ yang melekat pada nukleus leukosit nukleus polimorf.
Jumlah badan Barr dalam sel sama dengan jumlah total kromosom X dikurangi satu. Pada wanita normal dengan dua kromosom X jumlah tubuhnya adalah satu. Dalam sindrom triple X (XXX) jumlahnya meningkat menjadi dua; pada wanita dengan sindrom Turner yang hanya memiliki satu kromosom X (XO), tubuh Barr tidak ada. Pada laki-laki dengan sindrom Klinefelter yang memiliki kromosom XXY (trisomi), terdapat badan Barr.
Kehadiran kromosom Y pada laki-laki terdeteksi sebagai badan berfluoresensi intens (F-body) di dalam nukleus, ketika apusan bukal diwarnai dengan pewarna flurochrome dan diperiksa di bawah mikroskop fluoresensi. Karena teknik ini mahal dan slide cepat rusak, biasanya tidak digunakan untuk mempelajari status kromatin seks.
Struktur kimia Kromosom:
Pada analisis kimia setiap kromosom ditemukan mengandung DNA, sejumlah kecil RNA, protein histon dan non-histon, dan ion logam. DNA adalah konstituen molekul kromosom yang paling penting dan stabil.
Studi terbaru mengungkapkan bahwa setiap kromosom eukariota mengandung satu molekul DNA beruntai ganda terus menerus. Sebagian besar molekul DNA ada di kromosom sebagai struktur yang sangat melingkar atau terlipat. DNA dalam keadaan transkripsi aktif paling panjang dan menjadi eukromatik; Daerah DNA yang tidak aktif tetap sangat melingkar dan menjadi heterokromatik. Tingkat penggulungan DNA bervariasi dengan laju sintesis protein dalam fase siklus sel yang berbeda.
Dua jenis daerah heterokromatik permanen diamati pada kromosom manusia;
(a) Heterokromatin fakultatif mempengaruhi kromosom X yang tidak aktif pada wanita normal. Pada embriogenesis awal wanita, kedua kromosom X secara aktif terlibat dalam perkembangan ovarium; setelah itu salah satu kromosom X menjadi tidak aktif secara permanen dan membentuk badan Barr heterochromatic.
(b) Heterochromatin konstitutif diamati pada penyempitan kromosom primer dan sekunder. Urutan berulang basa DNA, kaya guanin dan sitosin, dikatakan hadir dalam heterokromatin konstitutif dan badan satelit. DNA berulang di beberapa bagian kromosom mungkin mengkode molekul intrinsik dalam bentuk RNA ribosom, RNA transfer, dan protein pengatur.
Histon adalah protein dasar yang kaya akan arginin dan lisin. Protein-protein ini terkumpul sebagai partikel sferoid di sepanjang untai DNA yang melingkari setiap partikel dan membentuk badan kompleks yang dikenal sebagai nukleosom atau badan-v (Gbr. 11-8). Setiap nukleosom terdiri dari empat pasang histon yang tersusun dalam dua kelompok simetris. Bukti eksperimental menunjukkan bahwa asosiasi DNA dengan nadanya menekan aktivitas gen.
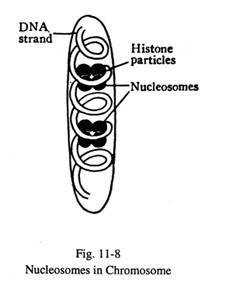
Protein non-histon bersifat asam dan membentuk banyak enzim, misalnya DNA polimerase dan RNA polimerase. Beberapa protein non-histon melepaskan histon dari aktivitas gen nukleosom dan derepres.
Prosedur analisis kromosom:
Untuk studi sitogenetik kromosom, dipilih sel yang tumbuh dan membelah dengan cepat dalam kultur. Jaringan yang paling umum digunakan adalah kulit, sumsum tulang, dan darah tepi.
Prinsip pembuatan kromosom dari darah tepi adalah sebagai berikut:
(a) Sekitar 1-2 ml. darah ditarik dari pembuluh darah, diheparinisasi dan diobati dengan phytohemagglutinin, diekstraksi dari kacang merah.
Phytohemagglutinin (PHA) merangsang limfosit (terutama sel-T) untuk berkembang biak dengan mitosis dan secara selektif memungkinkan aglutinasi dan sedimentasi eritrosit dewasa.
(b) Aliquot plasma dengan limfosit tersuspensi sekarang dipindahkan ke botol biakan dalam kondisi steril yang mengandung TC199 (Difco) sebagai media biakan. Inkubasi dalam botol kultur dilanjutkan selama kurang lebih 3 hari pada suhu 37°C dengan penambahan streptomisin dan penisilin sebagai pengawet.
(c) Colchicine sekarang ditambahkan ke kultur dan disimpan selama sekitar 2 jam. Colchicine menghentikan pembelahan sel pada metafase, dengan mencegah pembentukan mikrotubulus gelendong akromatik. Pada metafase, kromatid yang disatukan oleh sentromer berkontraksi secara maksimal.
(d) Sel dikumpulkan dengan sentrifugasi isi botol kultur. Larutan hipotonik natrium sitrat ditambahkan ke sel dan diinkubasi selama sekitar 20 menit. Larutan hipotonik memungkinkan sel membengkak dan menghilangkan kromosom.
(e) Media hipotonik dibuang dengan sentrifugasi. Sekarang fiksatif campuran etanol dan asam asetat ditambahkan ke pelet sel, dan dikocok perlahan untuk membentuk suspensi sel.
(f) Tetesan kecil suspensi sel ditempatkan di salah satu ujung slide yang dibersihkan secara kimiawi. Slide dibiarkan kering pada suhu kamar.
(g) Pewarnaan—Untuk studi pola kromosom konvensional, pewarnaan Giemsa banyak digunakan dengan hasil yang baik (Gbr. 11-9).
Identifikasi yang tepat dari kromosom individu sekarang dimungkinkan dengan mencatat pola pita pada kromosom setelah menerapkan salah satu dari empat teknik pewarnaan yang berbeda:
(i) Q-banding:
Ketika kromosom metafase tetap diwarnai dengan quinacrine hy drokloride atau quinacrine mustard, pita kromosom tertentu muncul sebagai daerah fluoresen di bawah mikroskop fluoresen. Pola pita-Q ini (neon) unik untuk setiap kromosom. Agaknya daerah pita-Q lebih kaya akan basis DNA adenin (A) dan timin (T) daripada daerah interband. Q-band yang sangat besar terlihat jelas di bagian distal lengan panjang kromosom Y, bahkan selama interfase.
(ii) G-banding:
Kromosom yang telah difiksasi diberi perlakuan ringan dengan enzim proteolitik (tripsin) sebelum pewarnaan. Enzim mampu mendenaturasi protein dalam kromosom. Ketika diwarnai dengan Giemsa setelah perawatan tersebut, pola pita-G pewarnaan gelap dapat terlihat pada kromosom di bawah mikroskop cahaya.
Daerah G-banding dan Q-banding kromosom berhubungan erat. Comings (1974) telah menyarankan bahwa protein yang tersisa setelah denaturasi dapat mencegah bahan pewarna masuk ke bagian DNA tertentu. Ada kemungkinan bahwa lebih sedikit protein dapat dikaitkan dengan DNA kaya AT; ini menjelaskan konkordansi G dan Q-band.
Pita G dan daerah Q-band adalah pasangan basa AT yang kaya; mereka berkorespondensi dengan daerah het erokromatin kromosom di mana replikasi DNA terjadi sedikit kemudian. Daerah interband kaya akan pasangan basa GC.
(iii) Pengikatan-R:
Ini adalah kebalikan dari G-binding, dimana daerah inter band ditunjukkan dengan pewarnaan Giemsa setelah pemanasan sampai 87°C. Pengikatan-R melengkapi pengikatan-G.
(iv) Pita-C:
Setelah perlakuan kasar terhadap kromosom tetap dengan alkali, asam atau garam, pewarnaan Giemsa mengungkapkan daerah yang diwarnai, pita С, dekat dengan sentromer. C-banding, bagaimanapun, tidak terlihat pada kromosom Y.
Banding kromosom membantu untuk menemukan kelainan tertentu dari struktur kromosom, seperti penghapusan dan translokasi daerah kromosom tertentu.
Kariotipe:
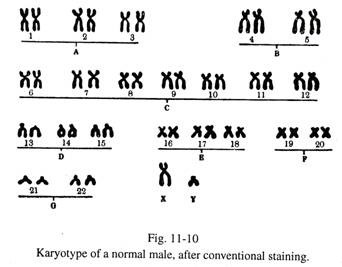
Ini adalah proses mengatur kromosom secara berurutan. Fotomikrograf yang diperbesar dari ‘ penyebaran’ kromosom yang diambil dari slide yang diwarnai. Kromosom individu dipotong dari foto, dicocokkan dengan pasangan homolog, dan disusun dalam urutan, kromosom terpanjang ditempatkan di awal dan terpendek di akhir.
Kromosom individu diidentifikasi menurut panjangnya, posisi sentromer, rasio panjang antara lengannya dan keberadaan badan satelit di lengannya. (Gbr. 11-10) Pola pita menambah identifikasi kromosom individu. (Gbr. 11-11).

Klasifikasi kromosom manusia:
Menurut klasifikasi ‘Sistem Denver’ (1960), kromosom manusia termasuk kromosom seks disusun menjadi tujuh kelompok dari A ke G, dalam urutan panjang yang menurun.
(1) Grup A:
Ini termasuk pasangan 1, 2, 3 kromosom. Masing-masing panjang dan metasentrik. Namun, kromosom 2 yang ditempatkan di Grup A adalah kromosom sub-metasentrik terpanjang .
(2) Grup Ð’:
Ini terdiri dari pasangan 4 dan 5 kromosom, yang cukup panjang dengan sentromer submetasentrik.
(3) Golongan С:
Ini adalah kelompok besar dan termasuk pasangan 6 sampai 12 kromosom; Kromosom X juga termasuk dalam kelompok ini. Kebanyakan dari mereka berukuran sedang dan sub-metasentrik. Pola pita membantu identifikasi kromosom individu.
(4) Grup D:
13 hingga 15 pasangan kromosom termasuk dalam kelompok ini. Semuanya berukuran sedang dan akrosentrik. Badan satelit dilekatkan pada ujung bebas dari lengan pendek setiap kromosom.
(5) Grup E:
Ini termasuk nomor kromosom 16 sampai 18. Mereka adalah kromosom submetasentrik yang cukup pendek.
(6) Grup F:
19 dan 20 kromosom berpasangan termasuk dalam kelompok ini. Masing-masing pendek dan metasentrik.
(7) Grup G:
Ini termasuk 21 dan 22 pasang kromosom; Kromosom Y termasuk dalam kelompok ini. Masing-masing sangat pendek dan akrosentris , 21 dan 22 kromosom menghadirkan badan satelit di lengan pendek mereka. Ujung distal lengan panjang kromosom Y menampilkan badan fluoresen setelah diwarnai dengan pewarna flurochrome.
Poin Pengamatan:
(a) 1 sampai 3 kromosom kelompok A, dan 19, 20 kromosom kelompok F bersifat metasentrik.
(b) 13 sampai 15 kromosom kelompok D, dan 21, 22 dan kromosom Y kelompok G bersifat akrosentrik. Lima pasang kromosom yang terdiri dari 13, 14, 15, 21, 22 memiliki badan satelit; karenanya disebut sat-kromosom. Sat-kromosom berkaitan dengan organisasi nukleolus.
(c) Kromosom lainnya bersifat submetasentrik.
Lokalisasi Gen pada Kromosom:
Lokalisasi gen pada kromosom manusia tertentu , meskipun sulit untuk ditentukan, dapat dinilai dengan analisis silsilah, dengan mempelajari pasien dengan penghapusan kromosom dan dengan mempelajari segregasi gen ‘penanda’ dalam keluarga dengan kelainan herediter tertentu. Gen penanda sering ditemukan pada populasi umum. Ciri-ciri penanda autosomal meliputi golongan darah dan protein serum tertentu.
Ciri-ciri penanda terkait-X meliputi buta warna, golongan darah Xg dan dalam beberapa kasus defisiensi dehidrogenase glukosa-6-fosfat. Studi silsilah telah menunjukkan hubungan yang erat antara lokus gen golongan darah ABO dan sindrom patela kuku, dan antara golongan darah Duffy dan salah satu bentuk katarak kongenital.
Pemetaan gen pada kromosom individu lebih ditingkatkan dengan menggunakan enzim restriksi (endonuklease) yang disintesis oleh banyak bakteri. Enzim restriksi membagi DNA menjadi fragmen-fragmen dengan panjang bervariasi dengan memotong di antara urutan spesifik basa, yang lokasinya berbeda untuk enzim yang berbeda. Polimorfisme panjang fragmen restriksi (RFLP) seperti itu bertindak sebagai sidik jari DNA, dan dideteksi dengan mengadopsi Teknologi DNA Rekombinan.
Analisis struktur DNA dengan RFLP, memungkinkan untuk menentukan induk mana yang menjadi sumber kromosom yang cacat. Ini membantu dalam konseling genetik, dalam penyelidikan kejahatan dan menentukan paternitas. Sekitar 50.000-100.000 gen diperkirakan terdapat dalam total genom manusia dengan 3 miliar pasangan basa. Pada awal 1993, lebih dari 2500 lokus telah ditempatkan pada posisi tertentu pada peta genetik manusia.
Abnormalitas sekitar 450 gen ini telah dikaitkan dengan penyakit manusia. Beberapa lokalisasi gen penting pada autosom ditempatkan bersama ini.
Kromosom:
1 – golongan darah Dufy, faktor Rh, protein histon, catract kongenital, retinitis pigmentosa.
2 – Asam fosfatase sel darah merah. Rantai ringan imunoglobulin Kappa.
5 – Hexosaminidase-B
6 – Kompleks histokompatibilitas utama (HLA), ataksia spino-cerebeller, sindrom adrenogenital.
7 – Gen struktural kolagen.
9 – Golongan darah ABO, sindrom kuku-patela.
14 – Rantai berat imunoglobulin
15 – Hexosaminidase-A 17- Timidin Kinase
19- Sensitivitas virus polio dan gema
20 – Adenosin deaminase
21 – Gen sindrom Down; gen untuk penyakit Alzheimer;
22 – Gen untuk rantai ringan lambda imunoglobulin
Kromosom X tampaknya mengandung lokus untuk dehidrogenase glukosa-6-fosfat, hemofilia A, penglihatan warna dan distrofi otot Becker pada lengan panjang, dan golongan darah Xg, ichthyosis vulgaris, albinisme okular, dan lokus keterbelakangan mental terkait-X. lengan pendek.
Kromosom Y mengandung gen determining pria ‘SRY’, komponen TDF (tesÂtis determining factor). Kehadiran satu kromosom Y menginduksi perkembangan testis; testis janin membebaskan testosteron dan faktor regresi mullerian, yang dengan aksi lokal memungkinkan diferensiasi tubulus dan duktus mesonefrik berkembang menjadi sistem duktus testis dan pada saat yang sama membantu regresi duktus paramesonefrik (sistem mullerian). Jadi kromosom Y melalui serangkaian peristiwa menginduksi perkembangan gonad jantan, saluran kelamin dan genitalia eksterna yang mengekspresikan fenotip jantan.
Namun pada sindrom ‘feminisasi testisâ €™ dengan kromosom XY, individu tampak sebagai perempuan sempurna dengan payudara dan alat kelamin luar perempuan, tetapi dengan testis intra-abdominal. Karena cacat genetik kromosom Y, sistem mullerian menjadi tidak responsif terhadap efek hormon laki-laki yang dibebaskan oleh testis janin.
Laki-laki normal memiliki susunan kromosom XY ; tetapi ketika seorang individu memiliki lebih dari satu kromosom X dengan kromosom Y tunggal (47, XXY; 48 XXXY), subjeknya adalah laki-laki dengan disgenesis tubulus seminiferus (sindrom Klinefelter). Oleh karena itu, kromosom Y menghadirkan gen penentu laki-laki yang kuat, terlepas dari jumlah kromosom X. Tetapi kehadiran kromosom X tambahan pada sindrom Klinefelter berdampak pada berkurangnya kesuburan dan membuat individu tersebut mengalami keterbelakangan mental.
Selain gen penentu laki-laki, kromosom Y mengandung gen hairy pinna dan antigen HY (histokompatibilitas). Panjang kromosom Y bervariasi dari orang ke orang dan mengikuti prinsip Mendelisme. Karena adanya antigen HY, cangkok jantan terkadang ditolak oleh betina dari strain yang sama.
Seorang wanita normal memiliki konstitusi kromosom XX. Pada embriogenesis awal, kedua kromosom X aktif secara genetik dan menginduksi perkembangan ovarium. Setelah itu, satu kromosom X menjadi heterochromatic dan ge netically inert, dan bertahan sebagai sex chromatin atau Barr body (Faculative heterochromatin). Ovarium janin tidak mengeluarkan hormon apa pun. Oleh karena itu dengan tidak adanya testis (dengan atau tanpa ovarium) sistem Wolffian (mesonephric) mengalami kemunduran dan sistem Mullerian (paramesonphric) berdiferensiasi menjadi organ seks wanita dan genitalia eksterna wanita.
Pada kesempatan langka individu dengan chromo XX beberapa konstitusi muncul laki-laki dalam fenotipe; ini menunjukkan adanya gen penentu testis di salah satu dari dua kromosom X yang berasal dari Y. Warisan langka ini dimungkinkan pada individu karena persilangan dalam gametogenesis di pihak ayah. Anehnya diamati bahwa orang dengan konstitusi kromosom 45, XO dapat tetap hidup, tetapi kombinasi 45, YO tidak dapat hidup.
Perubahan Struktur Kromosom (Gbr. 11-12):
Penghapusan:
Ini berarti hilangnya segmen kromosom, yang mungkin terminal atau interstitial. Penghapusan interstital yang dihasilkan dari dua patahan diikuti oleh penyatuan ujung yang patah. Pada sindrom ‘cri du chat’, bagian terminal dari lengan pendek kromosom 5 dihilangkan.
Translokasi:
Pertukaran segmen antara kromosom non-homolog dikenal sebagai translokasi. Proses translokasi membutuhkan pemutusan kedua kromosom non-homolog, diikuti dengan perbaikan yang mengarah ke susunan abnormal. Translokasi mungkin tidak selalu menghasilkan fenotipe abnormal, tetapi dapat menyebabkan pembentukan gamet yang tidak seimbang dan membawa risiko tinggi keturunan abnormal.
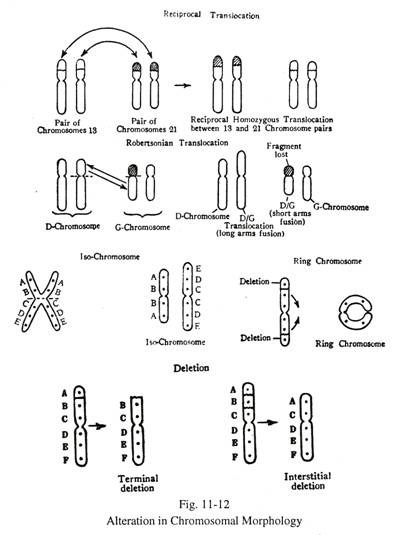
Translokasi resiprokal antara dua pasang kromosom non-homolog dapat menjadi heterozigot ketika hanya salah satu kromosom dalam pasangan yang terlibat, atau homozigot ketika kedua anggota pasangan kromosom bertukar segmen satu sama lain. Kadang-kadang translokasi melibatkan tiga pemutusan dan bagian kromosom yang rusak dimasukkan ke dalam kromosom non-homolog, sedangkan kromosom non-homolog lainnya menunjukkan penghapusan interstitial.
Translokasi Robertsonian atau fusi sentris adalah jenis translokasi khusus di mana pemutusan terjadi pada sentromer kedua kromosom dan seluruh lengan kromosom dipertukarkan. Pada pria, biasanya melibatkan dua kromosom akrosentrik, misalnya antara kelompok D dan G, 21/22 atau 21/21. Pada translokasi D/G, lengan panjang kromosom G melebur dengan lengan panjang kromosom D, dan fragmen yang dibentuk oleh fusi lengan pendek kedua kromosom hilang.
Ibu dari sindrom Down yang ditranslokasi biasanya merupakan pembawa translokasi D/G dengan hanya 45 kromosom. Dia menghasilkan empat jenis gamet — satu dengan kromosom D normal, satu dengan kromosom G normal, satu dengan kromosom D/G translokasi seperti ibu pembawa, dan satu dengan kromosom D/G dan kromosom G normal.
Keturunan yang berasal dari varietas gamet terakhir akan memiliki 46 kromosom tetapi akan menjadi trisomik untuk kromosom 21 dengan manifestasi sindrom Down. Oleh karena itu, ibu karier dengan translokasi D/G akan berisiko melahirkan anak dengan sindrom Down. Ketika seorang ibu mengalami translokasi yang melibatkan kedua kromosom 21, semua anaknya akan mengalami sindrom Down.
Pembalikan:
Bagian dari kromosom terlepas dan kemudian menyatu dengan kromosom yang sama dalam posisi terbalik. Gen tidak hilang tetapi ditempatkan di lokus yang diubah.
Iso-kromosom:
Sentromer kromosom, karena anafase abnormal (mitosis atau meiosis), membelah secara transversal, bukan membelah secara longitudinal. Hal ini berpuncak pada pembentukan dua kromosom dengan panjang yang tidak sama, masing-masing menghadirkan kromosom metasentrik dengan duplikasi gen. Kromosom yang dihasilkan berasal dari pembelahan transversal sentromer dikenal sebagai isokromosom.
Duplikasi:
Merupakan proses penambahan sebagian kromosom dari kromosom homolog lain dengan duplikasi gen. Efek duplikasi gen karena iso-pemisahan satu kromosom X kadang-kadang diamati pada sindrom Turner.
Kromosom cincin:
Kromosom cincin diamati ketika sebuah kromosom dihapus di kedua ujungnya, dan kemudian ujung ‘lengket’ yang dihapus direkatkan satu sama lain dalam bentuk cincin. Manifestasi kromosom cincin tergantung pada penghapusan gen tertentu.
Simbol yang Digunakan dalam Sitogenetik:
p—Lengan pendek kromosom
q—Lengan panjang kromosom
t—Translokasi; inv—Pembalikan
i—Iso-kromosom;
r—cincin kromosom
+or -Sign: Ketika ditempatkan sebelum simbol yang sesuai, itu berarti penambahan atau kehilangan seluruh kromosom. Misalnya, sindrom Down trisomi 21 dapat direpresentasikan sebagai 47, XY+21.
Ketika + atau – singns ditempatkan setelah simbol, ini menunjukkan peningkatan atau penurunan panjang kromosom. Sebagai contoh, sindrom cri du chat pada anak laki-laki dengan delesi lengan pendek kromosom 5 direpresentasikan sebagai 46, XY, 5p-
Pada kromosom Philadelphia atau Ph’ terjadi re ciprocal translocation antara lengan panjang kromosom 9 pita 34 dan lengan panjang kromosom 22 pita 11. Dengan demikian, kariotipe penyakit ini adalah-t (9; 22) (q34; ql 1).
Notasi selanjutnya disempurnakan untuk menentukan pita tertentu pada kromosom tertentu.
Garis diagonal melintasi kromosom atau jumlahnya menunjukkan mosaikisme, misalnya. XY/XX; XO/XX; XY/XXX; 45/46/47.
Gen:
Gen adalah unit hereditas dan terdiri dari bagian molekul DNA tertentu. Seperti disebutkan sebelumnya, gen disusun dalam rangkaian linier di dalam kromosom dengan urutan dan jumlah basis DNA yang tepat, berbeda untuk gen yang berbeda, dan memiliki awal dan akhir yang ditentukan. Karena kromosom tunggal mengandung satu heliks ganda molekul DNA dalam bentuk gulungan yang rapat, banyak gen atau sistron ditanggung oleh satu molekul DNA.
Posisi suatu gen dalam kromosom disebut lokus, yang diukur dengan mengacu pada sentromer. Biasanya gen tidak mengubah lokus, kecuali dalam rekombinasi selama persilangan atau dalam perubahan morfologi kromosom.
Gen yang menempati lokus identik pada sepasang kromosom homolog disebut alelomorf atau alel. Secara garis besar, gen alelik mengatur karakter fisik dan biokimia yang berbeda dari suatu individu. Dilihat dari tingkat molekuler, sepasang gen alelik mengatur sintesis satu rantai polipeptida.
Ketika gen alelik yang mengatur karakter atau sifat tertentu, katakanlah tinggi, bekerja ke arah yang sama (tinggi atau keduanya pendek), mereka disebut homozigot; ketika bekerja dalam arah yang berlawanan (satu tinggi dan lainnya pendek), alelnya heterozigot. Sebagian besar sifat herediter bersifat poligenik dan dihasilkan dari interaksi kompleks berbagai gen dan dipengaruhi oleh lingkungan. Terkadang, sepasang gen alelik dapat memengaruhi lebih dari satu karakter; ini dikenal sebagai pleiotropi.
Struktur kimiawi DNA (Gbr. 11-13):
Telah ditetapkan pada tahun 1953 oleh Wilkins, Watson, dan Crick pada difraksi sinar-X bahwa molekul DNA terdiri dari dua untai polinukleotida yang tersusun dalam heliks ganda. Setiap helai terdiri dari tulang punggung gula pentosa altenat (D-2-deoksiribosa) dan molekul fosfat, dan kedua helai disatukan oleh ikatan hidrogen antara basa nitrogen, yang melekat pada gula sebagai gugus samping dan mengarah ke pusat heliks.
Basa terdiri dari dua jenis, purin dan pirimidin. Purin dalam satu untai selalu berpasangan dengan pirimidin di untaian lainnya. Basa purin termasuk adenin (A) dan guanin (G); basa pirimidin termasuk timin (T) dan sitosin (C). Pasangan basa spesifik dalam kondisi normal (dalam bentuk keto) —adÂenin berpasangan dengan timin yang memiliki dua ikatan hidrogen dan dilambangkan dengan A =T; pasangan guanin dengan sitosin oleh tiga ikatan hidrogen dan diwakili oleh G=C.
Hal ini menunjukkan bahwa selama denaturasi DNA, pemisahan dua untai pada tingkat A=T lebih cepat dibandingkan dengan tingkat G=C. Namun, ketika basa dalam bentuk enol, adenin dapat berpasangan dengan sitosin dan guanin dengan timin. Ini adalah dasar dari mutasi gen.
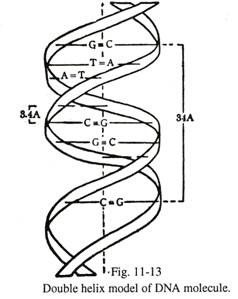
Dua untai molekul DNA saling melengkapi satu sama lain. Jika urutan basa dari satu untai diketahui, komposisi dasar dari untai lainnya dapat dirumuskan. Urutan basa dan jumlah nukleotida DNA bersifat spesifik, dan berbeda pada gen yang berbeda. Dengan demikian, bentuk DNA yang tak terhitung jumlahnya ada di dalam gen dan menyimpan beragam informasi genetik.
Fungsi molekul DNA:
Molekul DNA memiliki potensi berikut :
(1) Replikasi Diri
(2) Biosintesis RNA dan protein
(3) Rekombinasi;
(4) Mutasi.
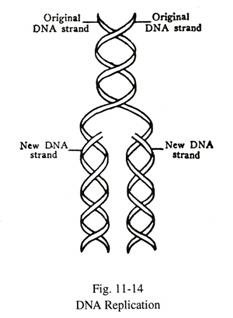
Replikasi Diri (Gbr. 11-14):
Selama pembelahan inti, dua untai molekul DNA terpisah, dan masing-masing untai bertindak sebagai templat dan mengatur pembentukan untai komplementer baru dari kumpulan nukleotida sebagai hasil pasangan basa spesifik. Dengan cara ini ketika sel membelah, informasi genetik ditransmisikan tidak berubah ke setiap sel anak. Kedua helai berpartisipasi dalam proses replikasi DNA, yang berlangsung pada fase-S (sintesis) siklus sel. Replikasi melibatkan beberapa enzim, seperti DNA polimerase, DNA ligase dan endonuklease spesifik.
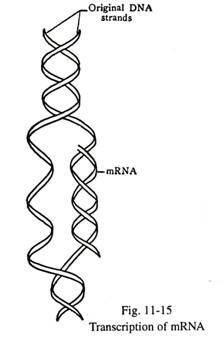
Biosintesis RNA dan Protein:
Molekul DNA juga bertindak sebagai templat untuk sintesis RNA, dan yang terakhir menyampaikan pesan genetik dan menguraikan sintesis rantai polipeptida spesifik protein melalui hubungan linier asam amino. Oleh karena itu, dogma sentral genetika molekuler mencakup DNA→RNA melalui proses transkripsi, dan protein RNA→ melalui translasi.
RNA (Asam nukleat ribosa) berbeda dari DNA pada dasarnya dalam tiga cara: ia biasanya memiliki rantai polinukleotida beruntai tunggal; gula pentosa adalah D-ribosa; dari empat basa organik, tiga mirip dengan DNA (Adenin, Guanin, Sitosin), dan yang keempat adalah urasil, bukan timin. Oleh karena itu, selama transkripsi dari DNA ke RNA, adenin berpasangan dengan urasil (A=U). RNA ada dalam tiga bentuk — messenger RNA (mRNA), ribosomal RNA (rRNA), dan transfer RNA (tRNA). Molekul DNA poligenik bertindak sebagai cetakan untuk ketiga varietas RNA. Tidak seperti replikasi DNA, hanya satu dari dua untai molekul DNA yang bertindak sebagai cetakan untuk RNA.
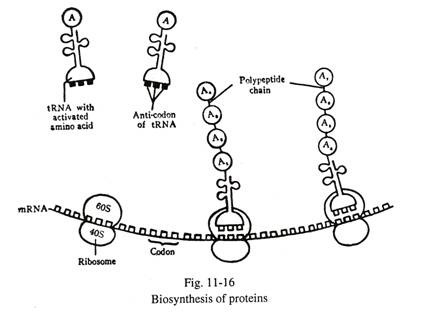
Rantai polinukleotida mRNA terbentuk di dalam nukleus di sisi salah satu untai molekul DNA dengan bantuan RNA polimerase. Selama sintesis RNA, dua untai DNA terpisah (Gbr. 11-15). Pemilihan untai DNA, untuk sintesis RNA, berlangsung dengan bantuan RNA polimerase I untuk rRNA, polimerase II untuk mRNA dan polimerase III untuk tRNA. Messenger RNA yang terbentuk menyampaikan pesan genetik dengan urutan dasar pelengkap, dan bergerak ke dalam sitoplasma melalui pori-pori inti.
Sejumlah ribosom sitoplasma (mengandung RNA ribosom dan protein) melekat pada rantai polinu kleotida mRNA. Ribosom adalah situs di mana rantai polipeptida protein dibentuk oleh hubungan linier asam amino yang berbeda.
Urutan dan nomor asam amino spesifik untuk protein yang berbeda; ini ditentukan oleh pembacaan yang tepat dari urutan dasar mRNA dalam arah ujung 5′ ke ujung 3′ . Dua puluh (20) asam amino terlibat dalam biosintesis protein. Sebelum pembentukan ikatan peptida, asam amino diaktivasi dan melekat pada salah satu ujung molekul RNA transfer (tRNA) spesifik. Sekuen basa tRNA yang membawa asam amino teraktivasi mengidentifikasi sekuen basa komplementer mRNA dan melekat pada mRNA terakhir dengan ikatan hidrogen sampai rantai polipeptida protein terbentuk.
Oleh karena itu mRNA, rRNA, tRNA dan sejumlah enzim secara aktif terlibat pada berbagai tahap biosintesis protein. Proses rumit biosintesis dari rantai polinukleotida mRNA ke rantai polipeptida protein dikenal sebagai translasi (Gbr. 11-16). Rantai polinukleotida mRNA dapat berupa monokistronik atau polikistronik.
Kode Genetik:
Karena basa DNA atau RNA dan asam amino protein disusun dalam urutan linier, pasti ada hubungan bersama antara basa nitrogen dan asam amino. DNA atau RNA menyajikan empat (4) basa, dan struktur primer protein terdiri dari dua puluh (20) asam amino. Setelah percobaan yang melelahkan Nirenberg dan Matthaei pada tahun 1961 menetapkan bahwa urutan tiga (3) basa mRNA (dan karenanya DNA komplementer) mengkode satu asam amino.
Karena tiga basa berurutan spesifik untuk satu asam amino, kemungkinan jumlah kombinasi dari empat basa yang diambil tiga basa sekaligus adalah 4 3 atau 64. Triplet berbasis nukleotida seperti itu disebut kodon. Akhirnya, semua 64 kodon ditemukan menentukan asam amino yang berbeda. Namun, tiga kodon seperti UAG, UGA, dan UAA tidak mengkode asam amino apa pun; karenanya ketiga ini disebut kodon non-sense atau terminal dan menandakan pemutusan rantai polipeptida.
Tiga basa tidak berpasangan yang melekat pada satu loop tRNA dikenal
sebagai anti-kodon yang cocok dengan kodon pelengkap mRNA. Sedangkan kodon dibaca dari ujung 5′ ke arah ujung 3′, antikodon dibaca dari arah 3′ ke 5′; seperti yang dinyatakan sebelumnya, tRNA membawa asam amino aktif di salah satu ujung rantai.

Kode Genetik pada mRNA, dan asam amino yang dikodekan.
Kodon mematuhi beberapa prinsip:
(a) Kodon tidak tumpang tindih dan mengikuti urutan yang ketat di sepanjang untai polinukleotida mRNA.
(b) Mereka bersifat universal dan berlaku untuk semua organisme.
(c) Kodon degeneratif – Ketika dua atau lebih kodon berdiri untuk asam amino yang sama, mereka dikatakan dalam bentuk degeneratif. Kode GUU, GUC, GUA, GUG untuk Valin; UUU, kode UUC untuk fenil alanin; UUA dan UUG singkatan dari leu cine. Dalam kebanyakan kasus, dua basa pertama tetap tidak terpengaruh dan perubahan basa ketiga menghasilkan degenerasi.
(d) Kodon yang ambigu atau salah menentukan asam amino yang berbeda. Dalam kondisi normal UUU singkatan dari fenil alanin, tetapi dengan adanya streptomy cin mungkin kode untuk leucine atau isoleuÂcine.
(e) Inisiasi atau kodon awal—kode AUG untuk metionin dan bertindak sebagai sinyal awal dalam sintesis rantai polipeptida. Urutan asam amino dalam rantai polipeptida dikenal sebagai struktur primer protein.
Gugus amino bebas di salah satu ujung rantai dikenal sebagai ujung N-terminal, dan gugus karboksil bebas di ujung lain rantai disebut ujung terminal С. Setiap asam amino dalam rantai disebut residu. Residu terminal N dianggap sebagai bilangan pertama, dan residu terminal-C sebagai bilangan terakhir dari urutan asam amino.
Metionin dalam kompleks inisiasi diformilasi oleh enzim spesifik sehingga ikatan peptida tidak terjadi pada ujung N-terminal. Dua kodon, AUG dan UGG hanya berdiri untuk satu asam amino; AUG untuk metionin dan UGG untuk triptofan.
(f) Kodon terminal atau non-sence. Tiga kodon seperti UAG, UGA, dan UAA tidak mengkode asam amino apa pun. Kodon terminal menandakan pemutusan rantai polipeptida.
Konsep organisasi gen saat ini:
- Seperti disebutkan sebelumnya, gen adalah bagian dari molekul DNA spesifik yang mengatur sintesis satu rantai polipeptida. Gen tipikal terdiri dari untaian DNA yang mencakup unit transkripsi dan wilayah promotor.
Unit transkripsi terdiri dari beberapa segmen ekson yang mendikte pembentukan protein, dipisahkan oleh segmen intron yang tidak diterjemahkan menjadi protein. PremRNA terbentuk dari DNA, dan kemudian intron dihilangkan dalam nukleus melalui proses penyambungan pasca transkripsi, sehingga mRNA akhir yang memasuki sitoplasma hanya terdiri dari ekson.
Daerah promotor terletak pada sisi ujung 5′ dari unit transkripsi gen. Ini berisi berbagai segmen DNA yang mendahului unit transkripsi dari ujung 3′ ke ujung 5′ dalam bentuk segmen Specifier, Quantifier dan Regulator. Urutan dasar dari segmen tertentu termasuk TATA (dikenal dengan TATA Box), yang memastikan bahwa transkripsi memulai titik yang tepat. Z-DNA merupakan segmen regio promotor, yang dapat menentukan ekspresi spesifik jaringan.
- Modifikasi pasca-translasi. Setelah rantai polipeptida diterjemahkan melalui mRNA, rRNA dan tRNA, produk protein akhir dimodifikasi oleh kombinasi reaksi yang meliputi hidroksilasi, karboksilasi, glikosilasi atau fosforilasi residu asam amino. Polipeptida yang lebih besar diubah menjadi bentuk yang lebih kecil melalui pemutusan ikatan peptida; setelah itu protein dilipat ke dalam konfigurasi kompleksnya.
Sel eukariotik tipikal mensintesis sekitar 10.000 protein berbeda selama masa hidupnya. Protein yang disintesis oleh gen dapat berupa salah satu dari tiga jenis—enzim, protein struktural, dan protein pengatur.
- Analisis sitogenetik pada mola hidatidosa, tumor atau trofoblas, menunjukkan bahwa ovum abnormal kehilangan nukleusnya sendiri dan dibuahi oleh dua sperma. Dengan demikian zigot mengandung dua pronuklei jantan, memiliki di antara mereka setidaknya satu kromosom X, Pada kehamilan mola lengkap membran trofoblas berkembang, tetapi embrio tidak muncul, Pencetakan genom menunjukkan bahwa kromosom ibu mengatur perkembangan embrioblas, dan kromosom paternal mengatur perkembangan trofoblas .
Rekombinasi:
Selama pindah silang pada meiosis, terjadi pertukaran materi genetik antara kromosom homolog. Hal ini menyebabkan rekombinasi atau pengocokan gen. Salah satu dari dua peristiwa dapat diamati dalam persilangan. Dua gen berbeda yang awalnya terletak pada kromosom yang sama dari pasangan kromosom tertentu, dapat dipisahkan satu sama lain dan selanjutnya didistribusikan ke kedua kromosom homolog; atau salah satu dari dua gen yang awalnya terletak di setiap kromosom homolog mungkin disatukan pada kromosom yang sama.
Ketika dua gen berbeda terletak pada pasangan kromosom yang sama, mereka dikatakan terkait. Pindah silang lebih mungkin terjadi antara gen pada kromosom tertentu yang berjauhan daripada gen yang berdekatan. Seseorang dapat menilai jarak relatif antara gen pada kromosom apa pun dengan menentukan frekuensi persilangan yang terjadi di antara gen-gen ini. Jarak genetik antara dua lokus pada kromosom tertentu dinyatakan dalam centimorgan (cM). Dua lokus berjarak 1cM, jika ada kemungkinan 1% persilangan di antara keduanya dalam meiosis. Rata-rata 30 sampai 35 persilangan per sel diperkirakan terjadi selama meiosis pada pria, dan mungkin dua kali lebih banyak selama meiosis pada wanita.
Dengan menentukan frekuensi rekombinasi akibat persilangan antar progeni, maka dimungkinkan untuk menyusun peta keterkaitan pada manusia dengan pengelompokan gen pada kromosom tertentu. (Vide supra, dalam lokalisasi gen pada kromosom)
Rekombinasi fragmen DNA dapat dipelajari secara eksperimental dengan memungkinkan fusi sel dari dua spesies berbeda dan kemudian menempatkannya dalam kultur. Hibrida sel yang menyatu mengandung konstitusi kromosom dari kedua spesies dan segmen pertukaran DNA saat mereka beregenerasi dan membelah. Semua proses regenerasi ini melibatkan pertukaran sekuens DNA secara acak, dan akhirnya sintesis protein berubah secara signifikan dari sel-sel leluhur yang telah melebur sebelumnya.
Pada tahun 1972, Jackson et al. menjelaskan metode biokimia untuk memotong molekul DNA dari dua organisme yang berbeda, menggunakan enzim restriksi, dan menggabungkan kembali fragmen untuk menghasilkan molekul DNA hibrida yang berfungsi secara biologis.
Selanjutnya, para ilmuwan berhasil sepenuhnya menyisipkan gen untuk kedua rantai insulin ke beberapa strain Escherichia Coli, dan setelah isolasi dan pemurnian, rantai A dan В bergabung dengan ikatan disulfida untuk menghasilkan insulin manusia. Dengan ditemukannya teknologi ‘DNA Rekombinan’, sejumlah zat esensial seperti insulin manusia, interferon, hormon pertumbuhan manusia, kalsitonin dan banyak lainnya diproduksi secara komersial.
Mutasi:
Perubahan pasangan basa molekul DNA dikenal dengan mutasi gen (mutasi titik). Karena gen bertanggung jawab untuk sintesis protein melalui transkripsi dari DNA ke RNA dan translasi dari RNA ke protein, mutasi mungkin memiliki efek beragam berikut pada protein yang sesuai:
(a) Kodon triplet yang diubah dapat mengkode asam amino yang sama tanpa ada perubahan pada protein yang dihasilkan. Sekitar 20 hingga 25% dari semua kemungkinan perubahan basis tunggal termasuk dalam tipe ini.
(b) Pada sekitar 70 sampai 75% kasus, mutasi basa tunggal dapat mengkode asam amino yang berbeda dan menghasilkan sintesis protein yang diubah yang menghasilkan penurunan atau kehilangan total aktivitas biologis.
(c) Dalam sekitar 2 sampai 4% kasus mutasi basa tunggal, triplet dapat menandakan pemutusan rantai peptida yang tidak mampu mempertahankan aktivitas biologis normal.
(d) Pada kesempatan langka, lebih dari satu basa tunggal dalam urutan DNA mungkin terlibat dalam mutasi gen. Akibatnya kadar enzim tertentu dapat berkurang karena tidak disintesis atau disintesis dengan aktivitas yang berkurang. Terkadang, mutasi gen dapat menyebabkan peningkatan sintesis enzim dengan aktivitas yang meningkat.
(e) Dalam beberapa kasus kelainan genetik, protein tertentu dapat disintesis, tetapi protein tersebut tetap tidak aktif secara fungsional. Hal ini terjadi pada sebagian besar kasus hemofilia.
Biasanya pasangan basa dalam replikasi atau transkripsi berlangsung dalam bentuk keto, di mana kombinasinya adalah A=T (dalam DNA), A=U (dalam RNA), G=C. Namun pada mutasi gen terjadi pasangan basa pada enolfrom, yang kombinasinya adalah A=C, G=T (pada DNA), G=U (pada RNA). Pasangan bair yang tidak biasa seperti itu dikenal sebagai tautomerisasi.
Mutasi dapat terjadi secara spontan atau diinduksi oleh berbagai zat kimia atau fisik, misalnya gas mustard, radiasi sinar-X, sinar gamma dari radium, dan atom radioaktif lainnya. Gen mutan dapat diwariskan atau muncul secara acak. Salah satu contoh khas mutasi gen diamati pada anemia sel sabit, di mana rantai beta hemoglobin dewasa yang mengandung 146 asam amino memiliki Valin, bukan asam glutamat, di posisi ke-6.
Sintesis DNA diarahkan RNA:
Telah disarankan oleh Temin pada tahun 1972, dari studi virus RNA bahwa aliran informasi genetik kadang-kadang terjadi dalam arah terbalik dari RNA ke DNA dengan bantuan reverse transcriptase. Virus semacam itu dikenal sebagai retrovirus, yang ketika dimasukkan ke dalam sel hewan inang bergabung dengan wilayah spesifik untaian DNA inti melalui proses rekombinasi.
Ini membentuk dasar dari studi tentang onkogen. Daerah DNA tertentu dalam sel normal berfungsi sebagai cetakan untuk sintesis RNA dan yang terakhir pada gilirannya bertindak sebagai cetakan untuk sintesis DNA, yang kemudian digabungkan dengan DNA inti. Amplifikasi yang dihasilkan dari daerah DNA tertentu membantu dalam diferensiasi embrio dan mungkin dalam patogenesis kanker.
Jenis Gen:
- Gen dominan mengekspresikan sifat fisik atau biokimianya, ketika gen alel bersifat homozigot atau heterozigot untuk sifat tersebut. Ini mengikuti pola pewarisan Mendelian dan dapat diamati dari catatan silsilah keluarga tersebut. Ketinggian disebabkan oleh gen dominan. Konstitusi genetik individu yang tinggi mungkin T:T atau T:t (T untuk tinggi, t untuk pendek). Sebagian besar sifat dominan diekspresikan dalam keadaan heterozigot (Gbr. 11-17).
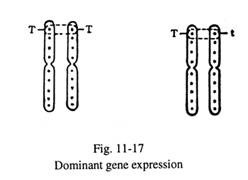
Kelainan genetik akibat mutasi gen autosomal dominan memiliki ciri- ciri sebagai berikut:
(a) Sifat tersebut diwariskan dari satu generasi ke generasi lainnya. Ini memiliki transmisi vertikal. Setiap orang yang terkena dampak biasanya memiliki orang tua yang terkena dampak. Terkadang kelainan itu bisa muncul tiba-tiba dalam satu generasi. Ini mungkin hasil dari mutasi baru; atau jika orang tua dengan gen abnormal meninggal pada awal kehidupan sebelum penyakit dapat bermanifestasi, riwayat kasih sayang orang tua mungkin kurang. Ini terjadi pada penyakit Huntington, di mana penyakit ini diekspresikan pada usia dewasa pertengahan.
(b) Bila salah satu orang tua terkena, risiko memiliki anak yang terkena adalah 50%.
(c) Karena sifatnya autosomal, kedua jenis kelamin mungkin sama-sama terpengaruh. Beberapa gen auto somal diekspresikan secara istimewa pada satu jenis kelamin. Ini disebut gen terbatas jenis kelamin. Asam urat dan kebotakan pra pikun lebih banyak menyerang laki-laki.
(d) Jika individu yang terkena dampak menikah dengan orang normal, setengah dari anak-anak mereka akan terpengaruh.
(e) Tingkat ekspresi sifat abnormal dapat berbeda pada anggota keluarga yang sama. Sebagai contoh, pada polidaktili beberapa anggota menunjukkan embel-embel seperti kutil kecil di sisi tangan, sedangkan anggota lainnya menunjukkan jari ekstra yang lengkap. Terkadang sebuah gen, ketika tidak menembus, mungkin tidak mengekspresikannya sama sekali. Jika seorang anak dan kakek nenek memiliki penyakit yang sama dan generasi tengah tidak menunjukkan manifestasi apapun, kondisi tersebut dikatakan telah melewati satu generasi.
(f) Anggota keluarga yang tidak terpengaruh tidak meneruskan sifat tersebut lebih lanjut.
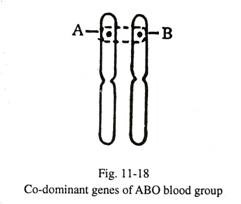
- Gen ko-dominan:
Ketika kedua gen alelik dominan tetapi dari dua jenis yang berbeda, kedua sifat tersebut dapat memiliki ekspresi bersamaan. Pada golongan darah ABO, gen A dan gen Ð’ keduanya dominan; ketika mereka menempati lokus identik dalam kromosom homolog, golongan darah AB diekspresikan (Gbr. 11-18).
- Gen resesif:
Mengekspresikan sifat hanya dalam keadaan homozigot yang berarti ketika kedua alel bersifat resesif untuk sifat tersebut (Gbr. 11-19). Oleh karena itu, mengikuti prinsip Mendel, susunan genetik individu pendek adalah t:t (t untuk pendek).

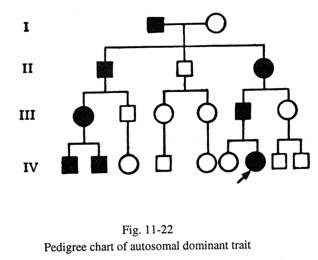
Penyakit yang disebabkan oleh mutasi gen resesif autosomal menunjukkan ciri-ciri sebagai berikut:
(a) Penyakit ini ditularkan oleh pasangan yang keduanya merupakan pembawa satu gen ab normal, tetapi mereka sendiri sehat karena alel lainnya normal.
(b) Pola penularan tampak mendatar pada analisis silsilah karena seringnya saudara kandung yang terkena, sedangkan orang tuanya normal.
(c) Risiko memiliki anak yang terkena dampak (dengan dosis ganda gen abnormal) pada pasangan karier adalah 25%. Oleh karena itu, sebagian besar pasangan pembawa, jika disarankan dengan benar, tidak akan mengambil risiko memiliki bayi lain yang terkena dampak kecuali fasilitas diagnostik prenatal tersedia untuk sifat tersebut.
(d) Sebagian besar kelainan metabolik diwariskan sebagai sifat resesif autosomal. Status heterozigot pasangan pembawa (memiliki satu anak yang terkena dampak) dapat dideteksi secara biokimia dalam beberapa kesalahan metabolisme bawaan. Tingkat enzim dalam heterozigot sekitar 50% lebih rendah dari kontrol.
(e) Karena kondisinya autosomal, kedua jenis kelamin cenderung sama terpengaruh.
(f) Orang tua dari individu yang terkena sifat resesif autosom sering berkerabat , karena perkawinan antara kerabat dekat (perkawinan sepupu) lebih cenderung membawa gen yang sama dari nenek moyang yang sama. Semakin jarang penyakit resesif, semakin besar frekuensi kekerabatan di antara orang tua dari individu yang terkena.
(g) Jika dua orang homozigot untuk kondisi resesif menikah dan memiliki anak, semua anak mereka akan terpengaruh. Tetapi tidak demikian dalam setiap kasus. Dalam satu keluarga kedua orang tuanya adalah albiÂnos (gangguan resesif), namun anak-anak mereka normal; pemeriksaan yang cermat terhadap sang ayah mengungkapkan bahwa dia memiliki jenis albinisme yang berbeda dari istrinya.
- Gen Pembawa:
Gen resesif heterozigot bertindak sebagai pembawa yang dapat diekspresikan pada generasi berikutnya. Ketika kedua orang tua heterozigot tinggi (T:t), kemungkinan tinggi anak adalah sedemikian rupa sehingga dari empat anak tiga tinggi dan satu pendek, dalam proporsi 3:1. Satu anak tinggi homozigot, dan dua lainnya heterozigot.
- Gen terkait seks:
Gen yang terletak pada kromosom X atau kromosom Y dikenal sebagai gen terpaut seks. Mutasi gen terkait-X lebih umum, dan sebagian besar dinyatakan sebagai sifat resesif.
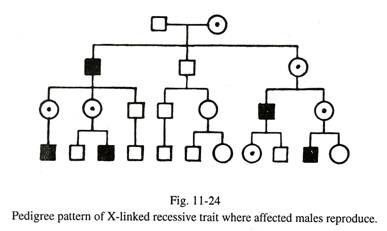
Sifat resesif terkait Х (Gbr. 11-20):
Hemofilia, buta warna parsial, defisiensi dehidrogenase glukosa-6-fosfat, distrofi otot Duchenne adalah contoh gen resesif mutan terkait-X. Ciri-ciri tersebut menunjukkan ciri-ciri sebagai berikut:
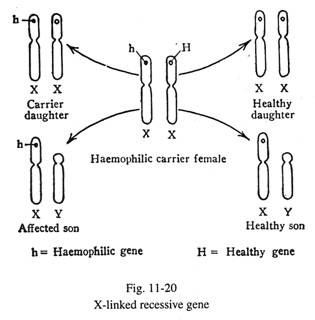
(a) Perempuan (XX) menjadi pembawa penyakit ketika satu kromosom X mengandung gen abnormal, sedangkan gen alelik dari kromosom X lainnya normal. Jadi betina tidak mengekspresikan penyakit dalam keadaan heterozigot. Di sisi lain, ketika gen abnormal melibatkan bagian non-homolog dari satu kromosom X laki-laki (XY), penyakit tersebut diekspresikan pada individu tersebut karena gen yang rusak tidak memiliki alel yang sesuai dalam kromosom Y untuk melawan. Oleh karena itu, laki-laki yang terkena disebut hemizigot. Secara garis besar, pada sifat resesif terkait-X, perempuan adalah pembawa dan laki-laki adalah korban dari penyakit tersebut.
(b) Bila ibu adalah karier dan ayah sehat, 50% anak laki-laki terkena penyakit dan 50% sisanya normal; 50% anak perempuannya adalah pembawa penyakit dan sisanya bebas. Oleh karena itu, ketika seorang anak laki-laki menderita hemofilik , ibunya harus menjadi pembawa dan 50% dari saudara perempuannya adalah pembawa penyakit tersebut. Namun, saudara laki-laki yang sehat atau saudara perempuan yang bebas pembawa dari individu penderita hemofilia tidak menularkan penyakit ini ke generasi berikutnya.
(c) Jika ibu adalah karier dan ayah menderita hemofilia, setengah dari anak laki-lakinya terkena dan setengahnya lagi sehat; setengah dari anak perempuan terpengaruh dan setengahnya adalah pembawa. Hal ini menunjukkan bahwa betina mungkin terpengaruh dalam kombinasi parental tersebut, tetapi kemungkinannya kecil karena jantan hemofilik biasanya meninggal lebih awal sebelum mencapai parenting. Kombinasi di atas lebih lanjut menunjukkan bahwa tidak ada penularan dari laki-laki ke laki-laki.
(d) Jika pejantan yang terkena tidak bereproduksi, pola silsilah dari sifat resesif terkait-X cenderung miring karena sifat tersebut ditransmisikan ke anak laki-laki dari saudara perempuan pembawa dari pejantan yang terkena.
(e) Pada kesempatan langka, seorang wanita mungkin menunjukkan sifat resesif terkait-X. Hal ini dapat dijelaskan sebagai berikut:
(i) Dia mungkin wanita Turner (XO);
(ii) Penampilan perempuan secara fisik disebabkan oleh feminisasi testis dengan kromosom XY;
(iii) Wanita yang terkena mungkin memiliki ibu pembawa dan ayah yang terkena; atau ibu karier dan ayah normal dengan mutasi baru yang memengaruhi kromosom X.
Sifat Dominan terkait-X:
Ini diamati pada rakhitis yang resisten terhadap vitamin D dan golongan darah Xg. Ciri -ciri sifat dominan adalah sebagai berikut:-
(a) Laki-laki yang terkena penyakit menularkan penyakit ke semua anak perempuannya, tetapi tidak kepada anak laki-lakinya.
(b) Baik laki-laki maupun perempuan terkena, tetapi penyakit ini tidak terlalu parah pada perempuan.
Warisan Y-Iinked:
Ini juga dikenal sebagai warisan holandric , di mana hanya laki-laki yang terpengaruh. Laki-laki yang terkena menularkan sifat itu kepada semua anak laki-lakinya, dan tidak kepada anak perempuannya. Penularan dari laki-laki ke laki-laki menunjukkan pewarisan terkait-Y.
Hairy pinna dan histokompatibilitas HY merupakan pewarisan holandrik manifes tigen.
Simbol yang digunakan dalam Bagan Silsilah (Gbr. 11-21):
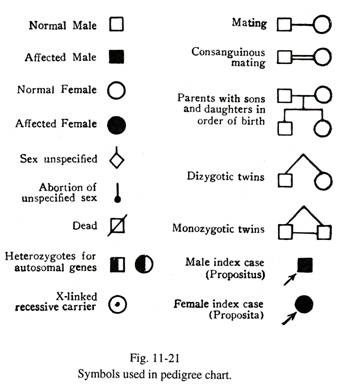
Warisan Autosomal Dominan (Gbr. 11- 22)
Beberapa contoh sifat dominan autosom—
saya. akondroplasia;
- Osteogenesis imperfekta;
aku aku aku. Brakidaktili, polidaktili, sindaktili;
- Pophyria sejati dengan urin port-wine karena porfirin;
- Keterbatasan jenis kelamin, asam urat dan kebotakan mempengaruhi laki-laki pra-pikun;
- Penyakit Huntington, muncul sekitar 50 tahun atau setelahnya;
- Edema angioneurotik;
viii. Hiperkolesterolemia familial;
- Diabetes insipidus;
- Sindrom Marfan, dimanifestasikan oleh ekstremitas memanjang, dislokasi lensa mata dan kelainan kardiovaskular;
- Sindrom patela kuku, dimanifestasikan oleh distrofi kuku, tidak adanya patela dan nefropati;
xii. Beberapa neurofibromatosis;
xiii. Kumparan poliposis.
Beberapa contoh sifat resesif terkait-X-
saya. Hemofilia-Hal ini disebabkan oleh globulin antihaemophilic yang rusak secara fungsional.
- Buta warna sebagian-Ini dinyatakan sebagai ketidakmampuan untuk membedakan antara merah dan hijau.
aku aku aku. Distrofi otot Duchenne.
- Defisiensi glukosa-6-fosfat dehidrogenase- Dimanifestasikan oleh anemia hemolitik ketika diobati dengan primaquin, phenacetin, nitrofurantoin, beberapa sulfonamid dan asam asetil salisilat .
- Feminisasi testis.
- Hunter’s syndrome-Hal ini disebabkan defisiensi enzim induronosulfate sulfatase, dan dimanifestasikan oleh ciri-ciri sindrom Hurler kecuali kekeruhan kornea.
Warisan Resesif Autosom (Gbr. 11- 22) 23):
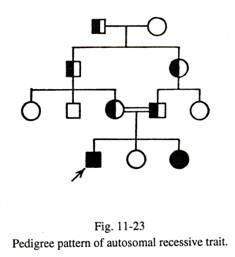
Beberapa Contoh sifat resesif autosom—
(1) Kesalahan metabolisme bawaan;
saya. Acatalasia, karena kekurangan enzim katalase; itu menyebabkan sepsis oral;
- Albinisme, depigmentasi kulit yang lengkap akibat defisiensi tirosinase;
aku aku aku. Alkaptonuria, di mana orang yang terkena mengeluarkan urin berwarna gelap karena adanya asam homogentisat. Hal ini disebabkan oleh kekurangan enzim homoge ntisic acid oxidase;
- Galaktosaemia, akibat defisiensi galaktosa-I-fosfat uridil transferase, dan dimanifestasikan dengan muntah dan diare akibat intoleransi terhadap galaktosa; ini diikuti oleh keterbelakangan mental, katarak dan sirosis hati;
- Sindrom Hurler, disebabkan oleh defisiensi enzim iduronidase, dan dimanifestasikan oleh keterbelakangan mental, kelainan tulang , hepatosplenomegali dan kornea berkabut.
- Phenylketonuria, karena kekurangan pheny lalanine hydroxylase dan bermanifestasi sebagai keterbelakangan mental, kulit peri dan epilepsi;
- Penyakit Tay-sachs, karena kekurangan hexosaminidase, dan bermanifestasi sebagai keterbelakangan mental, kebutaan dan kelainan neurologis.
(2) Hemoglobinopati:
saya. Pada anemia sel sabit, rantai beta mengandung valin di posisi ke-6, bukan asam glutamat. Ciri-ciri sel sabit heterozigot lebih tahan terhadap serangan malaria.
- Talasemia mayor diekspresikan pada homozigot, dan talasemia minor pada heterozigot.
(3) Imunoglobinopati:
saya. Beberapa gangguan imunologi mungkin disebabkan oleh sifat resesif autosomal.
Warisan Resesif Terkait-X (Gbr. 11-24):
Faktor Genetik pada Beberapa Penyakit Umum:
Diabetes mellitus:
Diabetes onset dini (juvenile-IDDM) lebih cenderung secara genetik daripada diabetes onset lambat. Beberapa peneliti berpendapat bahwa ia memiliki pewarisan resesif autosomal, sementara yang lain percaya bahwa ia memiliki warisan multifaktorial. Pada individu yang memiliki kecenderungan genetik, prediabetes dikenali dengan peningkatan kadar serum antibodi sel pulau.
Hipertensi esensial:
Ada dua aliran tentang cara pewarisan; satu sekolah menyatakan bahwa ia memiliki pewarisan multifaktor, sedangkan sekolah lain percaya bahwa itu karena mutasi gen dominan tunggal.
Penyakit jantung iskemik:
Penyakit jantung chaemic onset dini disebabkan oleh hiperkolesterolemia familial yang diwariskan sebagai sifat dominan autosomal. Pada sebagian besar individu yang terkena kondisinya bersifat multifaktorial dengan heritabilitas sekitar 65%.
Bisul perut:
Ulkus duodenum lebih sering terjadi pada individu dengan golongan darah О dan non-sekretor zat ABO. Empat puluh persen ulkus peptik memiliki kecenderungan turun-temurun.
Skizofrenia:
Ini diwariskan secara multifatorial dengan heritabilitas sekitar 85%. Beberapa percaya bahwa itu diwariskan sebagai sifat dominan au tosomal.
Beberapa Terminologi yang digunakan dalam Genetika
(1) Genom:
Genom menunjukkan set lengkap gen, haploid pada gama etes dan diploid pada sel somatik individu.
(2) Genotipe:
Ini berarti susunan genetik dari suatu individu yang ditetapkan pada saat pembuahan. Genotipe individu tinggi mungkin T:T (hoÂmozigot) atau T:t (heterozigot) yang dapat dinilai dengan analisis silsilah.
(3) Fenotip:
Ini berarti ekspresi fisik atau biokimia dari genotipe. Fenotipe berpotensi bervariasi dan merupakan hasil interaksi antara genotipe dan lingkungan tempat individu berkembang dan tumbuh. Mungkin saja individu dengan genotipe T:T bertubuh pendek. Ini mungkin karena beberapa gangguan endokrin atau nutrisi yang menekan aksi genotipe.
(4) Fenokopi:
Kadang-kadang perubahan lingkungan menghasilkan fenotipe baru, yang sangat mirip dengan penampilan yang disebabkan oleh genotipe tertentu. Bentuk fenotipe seperti itu dikenal sebagai fenokopi.
Melompat Gen atau Transposon:
Ini adalah kelompok elemen genetik yang benar-benar dapat berpindah dari satu tempat ke tempat lain dan dengan demikian memodifikasi atau menekan fungsi wilayah genetik target. Gen pelompat termasuk pseudogen, retrovirus dan onkogen, dan memiliki urutan DNA yang melompat. Setiap gen pelompat memiliki pengulangan basa terminal yang pendek dan serupa di kedua ujungnya.
Masing-masing memiliki sifat pengenalan urutan tertentu pada DNA target dan menghasilkan pengulangan langsung yang sama. Datang ke urutan target, gen bergerak menghasilkan istirahat asimetris pada untai berlawanan dari dupleks DNA dan kemudian terintegrasi di situs target.
Transposon mengontrol mutasi dan kombinasi ulang, dan mungkin bertanggung jawab atas amplifikasi gen. Status fungsional gen pelompat masih belum meyakinkan.
Konseling Genetik:
Setiap kali seseorang atau pasangan dengan kelainan genetik meminta saran, konselor genetik dihadapkan pada tiga masalah;
(a) Untuk menegakkan diagnosis yang tepat (genetik atau lingkungan) melalui pemeriksaan klinis dan pemeriksaan laboratorium ;
(b) Mendiskusikan prognosis dan nilai pengobatan yang mungkin dilakukan;
(c) Untuk menentukan risiko kekambuhan penyakit dalam keluarga dan menyelidiki deteksi pembawa, jika ada.
Studi kromosom dan kariotipe ditunjukkan dalam kondisi berikut;
(i) Pada bayi dengan kelainan kongenital yang melibatkan lebih dari satu sistem;
(ii) Dalam perkembangan seksual yang tidak normal;
(iii) Infertilitas, aborsi berulang, dll.
Ketika cacat kromosom (numerik atau struktural) terdeteksi dengan fenotipe abnormal , pengobatan, jika ada, bersifat simtomatik dan bukan kuratif.
Pembahasan tentang Risiko Kekambuhan dalam Keluarga:
(1) Jika kedua orang tua memiliki kromosom normal , meskipun anak dipengaruhi oleh kelainan kromosom (katakanlah, Trisomi 21-MonÂgol), orang tua dapat yakin bahwa kemungkinan terulangnya kondisi yang sama mempengaruhi anak di masa depan lebih kecil, karena penyebab kelainan ini adalah non-disjunction pada gametogenesis terutama yang melibatkan ibu tua, dan fenomena tersebut sebagian besar tidak disengaja.
Namun, jika kariotipe bayi Mongol menunjukkan translokasi antara kromosom G dan D (46) dan kariotipe ibu yang sehat menunjukkan kromosom translokasi yang seimbang, orang tua harus diberi tahu bahwa bayi Mongol yang serupa dapat berkembang lebih sering pada kehamilan berikutnya.
(2) Pada orang yang terkena dengan gen dominan autosomal heterozigot (katakanlah, Achondroplasia ), risiko kekambuhan di antara keturunannya adalah 1 banding 2 (50%), asalkan gen dominan tersebut sepenuhnya penetran.
(3) Pada gangguan resesif autosomal, ketika kedua orang tua sehat dengan gen resesif heterozigot untuk sifat yang sama, kemungkinan kambuh (katakanlah, fenilketonuria) di antara keturunannya adalah 1 sampai 4. Kita semua membawa sekitar 3 sampai 8 resesif merugikan gen, tetapi kemungkinan ekspresi gangguan resesif autosom jarang terjadi, kecuali dalam pernikahan kerabat. Ketika seorang ayah fenilketonurik menikah dengan sepupu pertamanya, peluang anak yang terkena adalah sekitar 1 banding 12, sedangkan dalam pernikahan dengan orang yang tidak terkait peluangnya adalah sekitar 1 banding 10.000.
(4) Dalam gangguan resesif terkait seks (katakanlah, hemofilia) ketika seorang anak laki-laki terpengaruh, ibunya yang sehat harus menjadi pembawa penyakit dan 50% dari saudara perempuannya adalah pembawa penyakit tersebut. Deteksi pembawa adalah tugas penting dari konselor genetik. Ketika laki-laki yang terkena terkait-X (katakanlah, buta warna sebagian) melahirkan anak, semua anak perempuan adalah pembawa dan semua anak laki-laki normal .
(5) Kadang-kadang individu mencari nasihat apakah ia akan terkena diabetes melitus, karena kedua orang tuanya menderita diabetes (gangguan resesif autosomal).
Dalam kejadian tersebut, selain dari analisis glukosa darah individu, titer antibodi sel anti-islet serumnya akan memberikan informasi apakah dia pra-diabetes. Dia kemudian disarankan untuk mengikuti pantangan makanan.
Deteksi Pembawa:
Operator dapat dideteksi dengan metode berikut:-
Tes biokimia:
(1) Katalase tingkat rendah pada acatalasia;
(2) Peningkatan kadar serum kreatin kinase pada distrofi otot Duchenne;
(3) Mengurangi faktor VIII pada hemofilia A;
(4) Pengurangan faktor IX pada hemofilia B;
(5) Penurunan glukosa-6-fosfat eritrosit pada defisiensi G-6-PD.
Amniosentasi:
(1) Penentuan jenis kelamin janin sebelum kelahiran dengan studi kromatin seks;
(2) rasio Lecithin-sphingomyelin cairan ketuban untuk mendeteksi maturitas paru janin;
(3) Tingkat alfa-fetoprotein cairan ketuban untuk deteksi anensefali dan spina bifida terbuka.
Footoscope:
Penggunaan foetoscope untuk mengumpulkan darah janin dari pembuluh darah umbilical membantu diagnosis prenatal anemia sel sabit dan beta thalassemia.



