Beberapa mode replikasi DNA yang paling penting adalah sebagai berikut!
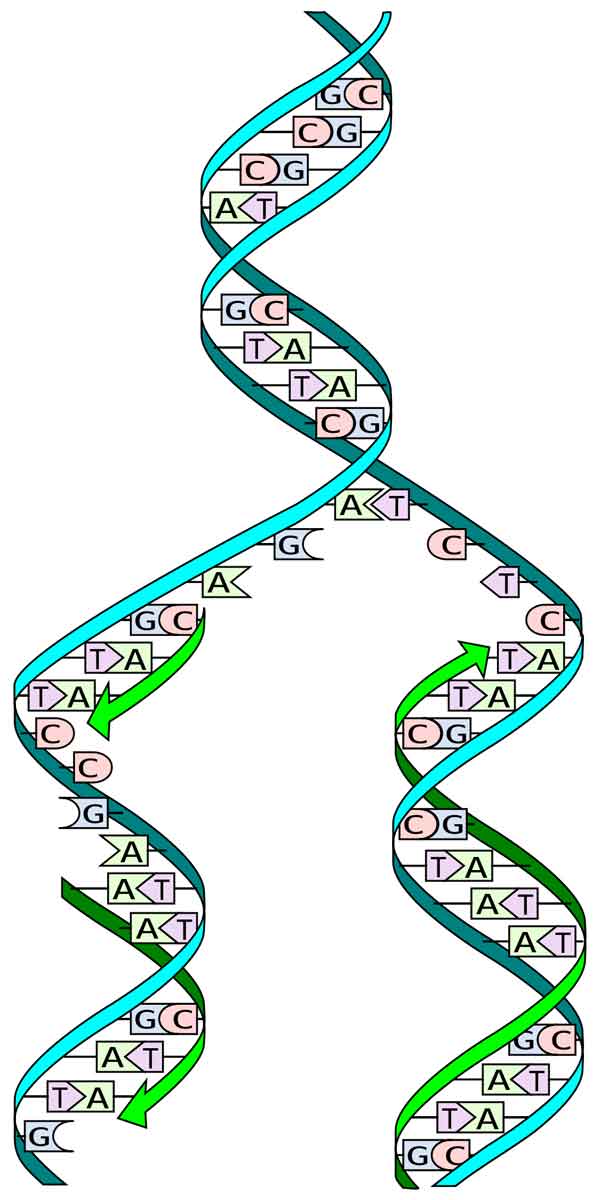
Replikasi DNA pada eukariota bersifat semikonservatif, semi-diskontinu, dan dua arah dibandingkan dengan semikonservatif, dua arah, dan kontinu pada prokariota.
Sumber Gambar: dbriers.com/tutorials/wp-content/uploads/2012/12/DNA_replication_split.png
Replikasi DNA terjadi selama fase-S siklus sel. Ini adalah proses kompleks multi langkah yang membutuhkan lebih dari selusin enzim dan faktor protein. Itu dimulai di tempat tertentu yang disebut asal replikasi atau ori. DNA bakteri dan virus memiliki asal replikasi tunggal. Ini berfungsi sebagai satu unit replikasi atau replika.
Dalam DNA eukariotik ada sejumlah asal replikasi. Ia memiliki beberapa segmen atau replika yang bereplikasi, yaitu multireplikanik. Tanpa ori, replikasi tidak akan terjadi. Persyaratan vektor untuk teknologi DNA rekombinan adalah untuk mendapatkan asal replikasi.
Replikasi DNA sangat mahal. Enzim utama replikasi DNA adalah DNA-dependent DNA polymerase. replikasi DNA cukup cepat. Replikasi DNA E. coli dengan 4,6 x 10 6 bp membutuhkan waktu 19 menit.
Rata-rata laju polimerisasi basa adalah 2000 bp per detik di setiap arah. Replikasi membutuhkan banyak energi yang berasal dari pemecahan trifosfat deoksiribonukleotida.
Replikasi terjadi sebagai berikut:
1. Aktivasi Deoksiribonukleotida:
deoksiribo atau deoksiribonukleosida monofosfat terjadi secara bebas di dalam nukleoplasma. Mereka terdiri dari empat jenis—deAMP (deoxyadenosine monophosphate), deGMP (deoxyguanosine monophosphate), deCMP (deoxycytidine monophosphate) dan deTMP (deoxythymidine monophosphate). Mereka pertama kali terfosforilasi dan berubah menjadi bentuk aktif yang memiliki tiga residu fosfat, bukan satu. Enzim fosforilase diperlukan bersama dengan energi.
Nukleotida terfosforilasi adalah deATP (deoxyadenosine triphosphate), deGTP (deoxyguanosine triphosphate), deCTP (deoxycytidine triph osphate) dan deTTP (deoxythymidine triphosphate). Trifosfat basa ini memiliki tujuan ganda. Mereka bertindak sebagai substrat serta menyediakan energi untuk polimerisasi nukleotida.
2. Paparan Untaian DNA:
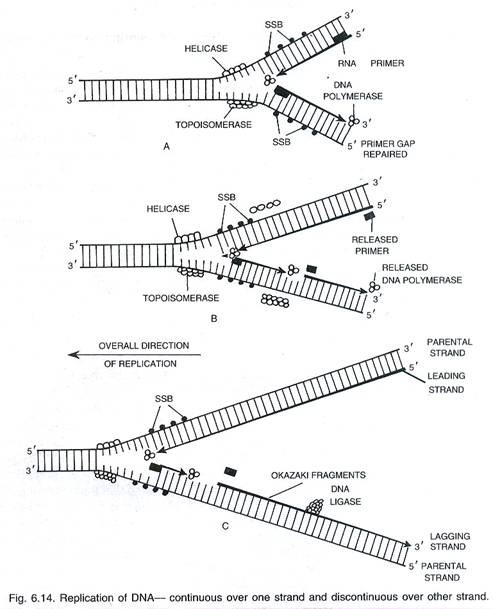
Enzyme helicase (unwindase) bekerja di atas situs Ori dan membuka ritsleting (melepaskan) dua untai DNA dengan menghancurkan ikatan hidrogen. Untaian yang terpisah distabilkan dengan menggunakan protein pengikat untai tunggal (SS BPs) atau protein penstabil heliks. Un winding menciptakan ketegangan di bagian uncoiled dengan membentuk lebih banyak supercoils. Ketegangan dilepaskan oleh enzim topoisomerase.
Mereka menyebabkan nicking dan resealing untai DNA. Seiring dengan topoisomerase, bakteri memiliki enzim lain yang disebut DNA gyrase yang dapat memperkenalkan superkoil negatif (pekerja yang lebih tua percaya bahwa gyrase berfungsi baik untuk helicase dan topoisomerase).
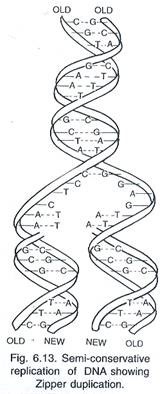
Dengan bantuan berbagai enzim, kedua untai DNA menjadi terbuka untuk replikasi. Namun, seluruh DNA tidak terbuka dalam satu bentangan karena kebutuhan energi yang sangat tinggi. Titik pemisahan berlangsung perlahan menuju kedua arah. Pada setiap arah, tampak struktur berbentuk Y yang disebut garpu replikasi (Gambar 6.13 dan 6.14).
3. Primer RNA:
Sangat penting untuk inisiasi rantai DNA baru. Primer RNA adalah untaian kecil RNA yang disintesis pada ujung 5′ untai DNA baru dengan bantuan enzim DNA polimerase RNA spesifik yang disebut primase. Primer RNA terbentuk pada ujung bebas satu untai dan ujung garpu untai lainnya. Pembentukan primer RNA merupakan fase inisiasi sintesis DNA karena tanpa adanya primer RNA, DNA polimerase tidak dapat menambahkan nukleotida.
Enzim yang lebih kompleks disebut primo some diperlukan dalam fag ф x 174 dan beberapa sistem prokariotik lainnya. Pada eukariota, fungsi primase dilakukan oleh enzim DNA polimerase α. Itu membangun ~ 10 basis RNA dan 20-30 basis DNA (Lewin, 2004). Setelah dimulainya rantai nukleotida, primer RNA dihilangkan dan celah diisi oleh DNA polimerase I pada prokariota dan DNA polimerase β pada eukariota.
4. Polimerase DNA:
Prokariota memiliki tiga jenis utama enzim sintesis DNA yang disebut DNA polimerase III, II dan I. Semuanya menambahkan nukleotida dalam arah 5’—>3′ pada 3′ —> 5′ bentangan untai induk. Mereka juga memiliki aktivitas exo-nuclease 3’—>5′. Sementara DNA polimerase III terutama terlibat dalam replikasi DNA (penambahan dan polimerisasi basa baru), polimerase I adalah enzim perbaikan utama. Polymerase II adalah enzim perbaikan kecil.
DNA polimerase I juga memiliki aktivitas eksonuklease 5—>3. Pada eukariota ditemukan lima jenis DNA polimerase— α, β, γ, δ, dan ε, tetapi tiga yang utama adalah α, δ dan e. Polymerase 8 terlibat dalam replikasi untai terdepan. Polymerase dapat membantu dalam sintesis lagging strand bersama dengan peran lainnya. Polymerase α adalah enzim terbesar dan utama dalam replikasi DNA. Semua DNA polimerase memiliki konfigurasi tangan yang mencengkeram dengan ibu jari di satu sisi, jari di sisi lain dan situs katalitik cekung seperti telapak tangan untuk menggabungkan templat dan pasangan basa.
5. Pasangan Basis:
Dua untai DNA yang terpisah dalam garpu replikasi berfungsi sebagai templat. Deoxyribonucleoside triphosphates berada di seberang basa nitrogen dari cetakan DNA yang terbuka — deTTP di seberang A, deCTP di seberang G, deATP di seberang T dan deGTP di seberang C.
Serangan nukleofilik memisahkan pirofosfat (PPi) dari triphosfat . Keterkaitan fosfodiester terbentuk. Hidrolisis pirofosfat oleh enzim pirofosfatase melepaskan energi. Deoxyribouncleoside triphosphate → Deoxyribouncleoside monophosphate + PPi
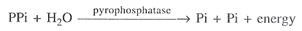
Energi tersebut digunakan untuk membentuk ikatan hidrogen antara nukleotida bebas dan basa nitrogen dari cetakan.
6. Formasi Rantai:
Ini membutuhkan DNA polimerase III (Kornberg, 1956) pada prokariota dan polimerase δ /ε pada eukariota. DNA polimerase III adalah enzim kompleks yang memiliki tujuh subunit (a, β, Æ , Æ´, €, θ, Ï„). Di hadapan Mg 2+ , ATP (GTP), TPP dan DNA polimerase III, nukleotida yang berdekatan ditemukan melekat pada basa nitrogen dari setiap untai DNA templat membentuk ikatan fosfodiester dan dihubungkan untuk membentuk untai DNA yang direplikasi.
Saat replikasi berlangsung, area baru dupleks DNA induk terlepas dan terpisah sehingga replikasi berlangsung cepat dari tempat asal menuju ujung lainnya. Primer RNA dihilangkan dan celah diisi dengan nukleotida komplementer melalui DNA polimerase I. Karena pembukaan rantai ganda DNA secara berurutan dan replikasinya membentuk dua rantai, replikasi DNA juga disebut duplikasi ritsleting.
Namun, DNA-polimerase dapat mempolimerisasi nukleotida hanya dalam arah 5’→ 3′ pada untai 3′ —> 5′ karena menambahkannya pada ujung 3′. Karena dua untai DNA berjalan dalam arah yang antiparalel, kedua templat tersebut memberikan ujung yang berbeda untuk replikasi. Replikasi atas dua templat dengan demikian berlangsung dalam arah yang berlawanan. Satu untai dengan polaritas У —> 5′ membentuk untai komplementernya secara terus-menerus karena ujung 3′ ujung terakhir selalu terbuka untuk pemanjangan.
Ini disebut untaian terdepan. Replikasi terputus-putus pada templat lain dengan polaritas 5′ → 3 karena hanya segmen pendek untai DNA yang dapat dibangun dalam arah 5′ → 3 karena pemaparan bentangan kecil templat pada satu waktu. Segmen pendek DNA yang direplikasi disebut fragmen Okazaki (= segmen Okasaki; Reiji Okazaki, 1968). Masing-masing memiliki 1000-2000 bp pada prokariota dan 100-200 bp pada eukariota .
Primer RNA juga diperlukan setiap kali fragmen Okazaki baru dibuat. Setelah mengganti primer RNA dengan deoksiribonukleotida dan polimerisasinya, fragmen Okazaki bergabung bersama melalui enzim, DNA ligase (Khorana, 1967). Untai DNA yang dibangun dari fragmen Okazaki disebut untai lagging.
Saat satu untai tumbuh terus menerus sementara untaian lainnya terbentuk secara terputus-putus, replikasi DNA bersifat semi-terputus. Karena replikasi berlangsung dua arah dari asal replikasi atau ori, satu untai induk akan membentuk untai terdepan di satu sisi dan untai tertinggal di sisi lain. Kebalikannya terjadi pada untaian induk di sisi lain. Ini membantu dalam menyelesaikan replikasi secara bersamaan di seluruh replika.
7. Proof-reading dan Perbaikan DNA:
Basis yang salah kadang-kadang diperkenalkan selama replikasi. Frekuensinya satu dari sepuluh ribu. DNA polimerase III mampu merasakan hal yang sama. Ini mundur, menghapus basis yang salah, memungkinkan penambahan basis yang tepat dan kemudian melanjutkan. Namun, bahkan DNA polimerase III tidak dapat membedakan urasil dari timin sehingga sering dimasukkan sebagai pengganti timin. Ketidakcocokan seperti itu dikoreksi melalui sejumlah enzim.
Ada mekanisme perbaikan terpisah untuk setiap kerusakan yang disebabkan DNA karena mutasi, paparan sinar UV atau ketidaksesuaian yang lolos dari mekanisme proof-reading. Sobekan atau kerusakan disebabkan oleh endonuklease di dekat daerah perbaikan. DNA polimerase I (Komberg, 1969) menghilangkan nukleotida yang tidak cocok atau salah jika ada dan mensintesis pengganti yang benar dengan menggunakan untaian utuh sebagai templat. Segmen yang baru terbentuk disegel oleh DNA ligase.



