Inilah catatan berguna Anda tentang Anthocerotopsida!
Kelas Anthocerotopsida (Anthocerotae) terdiri dari satu ordo, Anthocerotales dan satu famili, Anthocerotaceae, 6 genera dan 301 spesies. Menurut Muller (1940), Reimers (1954) dan Proskauer ordo Anthocerotales meliputi dua famili, (i) Anthocerotaceae dan (ii) Notothylaceae. Yang terakhir termasuk genus tunggal, yaitu Notothylas. Namun, menurut ahli briologi papan atas, hanya ada satu famili, yaitu Anthocerotaceae. Sekitar lima atau enam genera termasuk dalam keluarga ini.

Sumber Gambar: upload.wikimedia.org/wikipedia/commons/c/ce/Phaeoceros_laevis.jpg
Genera ini adalah—Anthoceros, Phaeoceros, Aspiromitus, Notothylas, Dendroceros dan Megaceros. Empat genera diakui secara universal, yaitu—Anthoceros, Megaceros, Dendroceros, dan Notothylas. Kelompok ini berbeda dalam banyak hal dari Bryophyta lainnya.
Namun, kelompok tersebut ditempatkan di antara Hepaticopsida (Hepaticae) dan Bryopsida (Musci). Kelompok ini dianggap sangat penting dari sudut pandang morfologinya, karena posisinya berada di antara dua kelompok penting, yaitu Hepaticopsida dan Bryopsida.
Ciri-ciri yang paling khas dari kelompok ini adalah sebagai berikut: Tubuh tumbuhan gametofit adalah thalloid dan dorsiventral. Rizoidnya sederhana dan berdinding halus. Rizoid tuberkulosis dan sisik ventral sama sekali tidak ada.
Jaringan thallus tidak terdiferensiasi. Ruang udara dan pori udara tidak ada. Setiap sel talus memiliki kloroplas besar dan pirenoid yang mencolok di dalamnya.
Antheridia bersifat endogen, yaitu muncul dari sel-sel hipodermis thallus di sisi dorsalnya. Antheridia dikembangkan di dalam ruang antheridial, sendiri-sendiri atau berkelompok di sisi dorsal thallus.
Archegonia ditemukan dalam kondisi cekung pada sisi dorsal thallus.
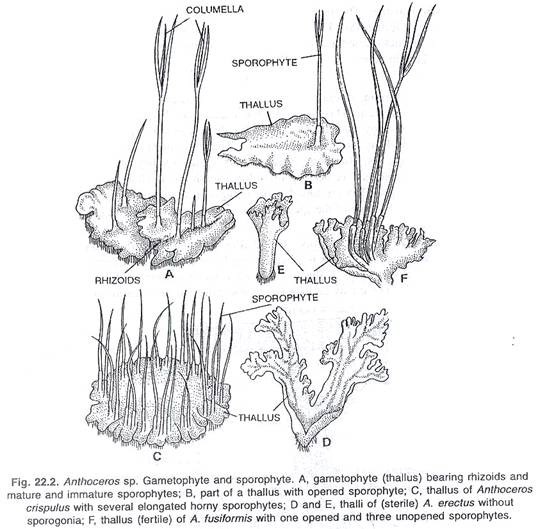
Sporogonium muncul dari sisi dorsal thallus. Strukturnya memanjang dan silindris. Ini terdiri dari kaki, daerah meristematik dan kapsul. Ini memiliki meristem kabisat, dan terus tumbuh sepanjang musim tanam. Dinding sporogonia mengandung klorofil. Bagian sentral steril adalah columella, yang dikelilingi oleh jaringan sporogenous dan spora. Para elater juga hadir.
Massa sporogenous berkembang dari amphithecium dan lengkungan di atas columella.
Keluarga-Anthocerotaceae:
Ada lima genera (yaitu, Anthoceros, Phaeoceros, Aspiromitus, Dendroceros dan Megaceros) dalam keluarga ini.
Fitur karakteristik:
Sporogonium (kapsul) adalah linier dan vertikal. Umumnya stomata terdapat pada dinding kapsul. Archesporium muncul dari amphithecium. Elaters bersel empat, halus atau berdinding tebal dan dengan atau tanpa pita penebalan. Genus Anthoceros telah dibahas di sini secara rinci.
Genus ANTHOCEROS:
Habitat dan distribusi:
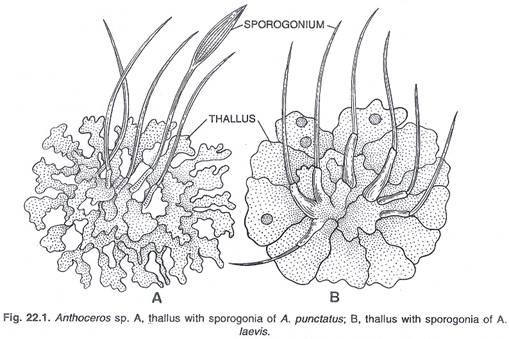
Sekitar 200 spesies dari genus ini ditemukan di seluruh dunia di daerah beriklim sedang dan tropis. Spesies ini lembab dan menyukai keteduhan. Sekitar 25 spesies telah dilaporkan dari India, oleh berbagai pekerja. Tiga spesies umum Himalaya adalah, Anthoceros himalayensis, A. erectus dan A. chambensis.
Spesies yang disebutkan di atas umum di berbagai daerah perbukitan seperti, Mussoorie, perbukitan Kumaon, lembah Chamba dan tempat lainnya, 5.000 kaki hingga 8.000 kaki. Beberapa spesies telah dilaporkan dari India Selatan. Semua spesies ditemukan di tempat yang sangat teduh dan lembab. Mereka ditemukan di cekungan bebatuan lembab di tambalan padat. Menurut Cavers (1911) beberapa spesies tumbuh pada kayu yang membusuk. Spesies ini, sama sekali tidak tahan kekeringan.
Struktur eksternal thallus:
Thallus kecil, sujud, hijau tua dan dibedakan secara dorsiventral. Thallus adalah lobus dan lobus agak terbagi. Tulang rusuk tengah tidak ditemukan. Permukaan dorsal talus A. laevis halus, sedangkan beludru seperti pada A. crispulus (Bhardwaj, 1950) atau mungkin dengan duri dan tonjolan seperti pada A. fusiformis.
Dalam setiap kasus ditemukan rizoid sederhana berdinding halus. Sisik ventral dan rizoid tuberkulat sama sekali tidak ada. Thalli berwarna hijau tua, karena adanya koloni Nostoc, yang dapat dengan mudah dilihat dengan bantuan lensa dari bagian bawah thallus.
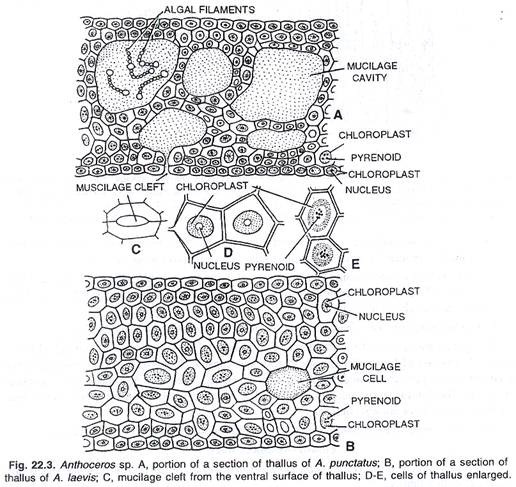
Struktur internal thallus:
Anatomi thallus cukup sederhana. Di dalam epidermis atas dan bawah terdapat sel-sel parenkim sederhana. Sel-sel parenkim adalah isodiametrik dan seragam. Ruang udara dan pori udara tidak ada. Setiap sel mengandung kloroplas besar yang memiliki pirenoid tunggal di tengahnya. Kloroplas berbentuk lensa. Kloroplas sel superfisial lebih panjang dari kloroplas sel lain.
Menurut McAllister (1914, 1927) pyrenoids dari Anthoceros sangat berbeda dalam struktur dari Chlorophyceae. Di sisi ventral thallus ditemukan beberapa rongga lendir antar sel. Rongga-rongga ini terbuka melalui bukaan kecil, yaitu pori-pori lendir di permukaan ventral thallus.
Kadang-kadang, koloni alga hijau biru bentuk-Nostoc, ditemukan di rongga lendir. Tidak ada hubungan simbiosis antara Nostoc, koloni, dan thalli ini. Menurut Peirce (1960) koloni-koloni ini, bagaimanapun, membahayakan thalli. Nukleus terletak di sekitar kloroplas, dekat pirenoid. Terkadang kloroplas membungkus nukleus di dalamnya.
Pertumbuhan apikal:
Pertumbuhan apikal Anthoceros adalah topik yang kontroversial, apakah itu terjadi oleh satu sel apikal atau sekelompok sel apikal. Menurut Smith (1955) pertumbuhan apikal talus diprakarsai oleh satu sel apikal.
Menurut Leitgeb (1879) pertumbuhan apikal pada genus ini terjadi oleh beberapa sel apikal marjinal. Menurut Mehra dan Handoo (1953) pertumbuhan apikal pada Anthoceros erectus dan A. himalayensis terjadi oleh sekelompok sel yang terletak pada depresi di puncak thallus.
Reproduksi:
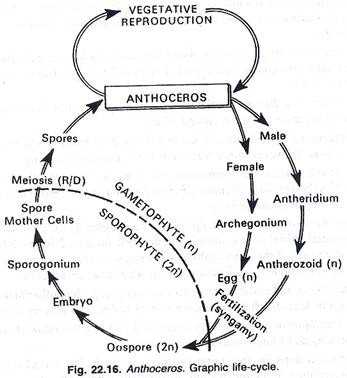
Reproduksi dilakukan dengan cara (1) vegetatif dan (2) seksual.
- Reproduksi vegetatif:
Reproduksi vegetatif terjadi dengan berbagai cara.
(a) Dengan pertumbuhan progresif dan kematian thallus:
Perbanyakan vegetatif terjadi dengan pertumbuhan progresif dan kematian bagian yang lebih tua dari thallus yang mencapai dikotomi. Tetapi metode ini tidak begitu umum di Anthoceros seperti di Riccia dan Marchantia.
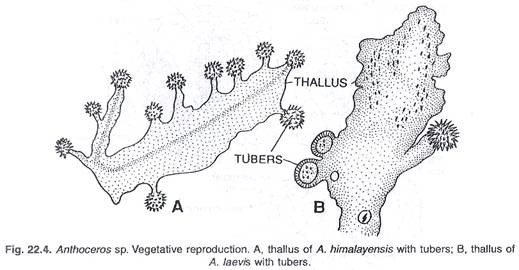
(b) Dengan umbi:
Pada spesies Anthoceros tertentu, talus menjadi menebal di beberapa tempat di pinggiran. Penebalan marjinal seperti itu disebut umbi. Umbi ini adalah struktur perennating. Mereka bertahan hidup dalam kondisi kekeringan. Pada munculnya kondisi yang menguntungkan, mereka berkembang menjadi thalli baru. Umbi terbentuk pada A. laevis, A, tuberosus, A hallii, A. pearsoni dan A. himalayensis.
(c) Oleh gemmae:
Pada beberapa spesies Anthoceros, gemmae telah ditemukan. Gemmae telah dicatat dari spesies, A. glandulosus, A. formosae, dll. Setiap gemma tersebut berkembang menjadi thallus baru.
(d) Dengan apeks yang tumbuh terus-menerus:
Menurut Campbell, thalli dari spesies A. pearsoni dan A. fusiformis menjadi benar-benar kering di musim panas, meninggalkan apeks yang tumbuh dengan jaringan yang berdekatan. Apeks ini menghadapi kondisi kekeringan. Pada pendekatan kondisi yang menguntungkan, apeks ini berkembang menjadi thalli baru.
- Reproduksi seksual:
Spesies Anthoceros mungkin homothallic (berumah satu) atau heterothallic (dioecious). Beberapa spesies homothallic adalah- A. fusiformis, A. punctatus, Kashyap (1915, 1929) telah mencatat, A. himalayensis sebagai spesies dioecious (heterothallic), tetapi Mehra dan Handoo (1953) telah melaporkan hal yang sama sebagai monoecious (homothallic). ) tapi protandrous. Spesies heterothallic adalah, A. pearsoni, A. halli, A. erectus dan lain-lain. Organ seks, yaitu, antheridia dan archegonia ditemukan tertanam dalam jaringan sisi dorsal thallus.
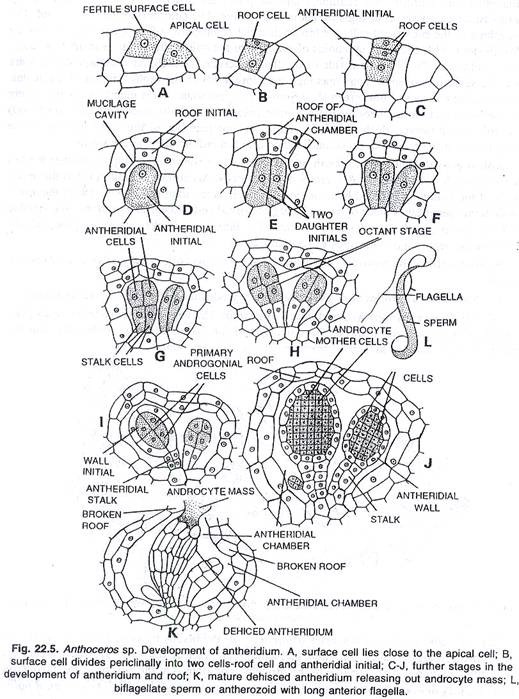
Perkembangan anteridium:
Antheridia diproduksi sendiri-sendiri atau berkelompok di ruang antheridial. Perkembangannya bersifat endogen. Meskipun antheridium berkembang dari sel superfisial, namun ia tertutup di dalam ruang antheridial yang tidak terbuka oleh lubang apa pun.
Sel superfisial dorsal thallus, terletak di dekat apeks yang tumbuh membelah secara periklin sehingga menghasilkan dua sel anak. Menurut Cavers, Campbell dan Haupt, sel superfisial membelah secara melintang dan tidak periklin. Sel anak atas bertindak sebagai inisial atap dan yang lebih rendah sebagai inisial antheridial.
Akhirnya, ruang berisi lendir muncul di antara inisial atap dan inisial antheridial. Rongga lendir ini membesar dan akhirnya menjadi ruang antheridial. Atap awal tidak ada kaitannya dengan perkembangan pada anteridium.
Ini membelah dan membelah kembali beberapa kali secara antiklin dan periklin sehingga menimbulkan atap dua lapis ruang antheridial. Secara bersamaan, inisial antheridial berkembang menjadi antheridium tunggal atau dalam kelompok antheridia. Sebuah antheridium tunggal berkembang di A. pearsoni dan terkadang di A. himalayensis.
Menurut Mehra dan Handoo (1953), pada A. erectus sejumlah antheridia berkembang dalam ruang antheridial. Di sini, awal antheridial membelah berkali-kali secara antiklin menghasilkan banyak sel dan setiap sel yang dihasilkan, berkembang menjadi antheridium. Perkembangan selanjutnya adalah sebagai berikut:
Inisial antheridial membelah dua kali dengan dinding vertikal yang berpotongan satu sama lain di sudut kanan, sehingga menimbulkan empat sel. Ini diikuti oleh pembagian melintang lainnya yang menghasilkan dua tingkatan yang masing-masing terdiri dari empat sel. Empat sel dari tingkat atas membelah secara melintang sehingga menimbulkan delapan sel, tahap oktan.
Semua sel tahap oktan membelah secara periklin menghasilkan delapan sel jaket primer luar dan delapan sel androgonial primer dalam. Empat sel dari tingkat yang lebih rendah berkembang menjadi tangkai anteridium yang terdiri dari empat baris sel.
Sel-sel androgonial yang terbentuk membelah berulang kali. Generasi terakhir dari sel-sel ini adalah sel induk androsit. Menurut Bagchee (1924) setiap sel induk androsit membelah secara diagonal menghasilkan dua androsit. Setiap androsit bermetamorfosis menjadi gelendong seperti biciliate antherozoid.
Struktur anteridium dewasa dan dehisensinya:
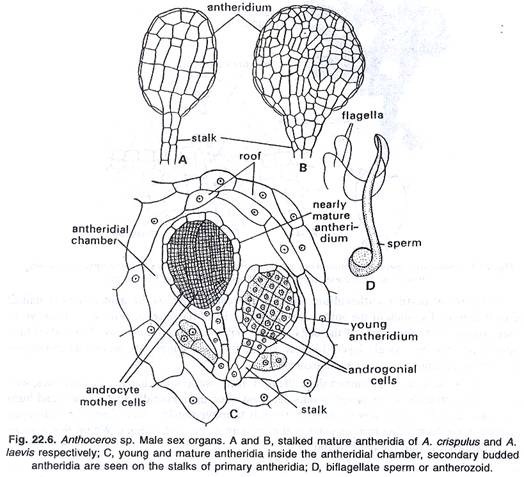
Anteridium dewasa bertangkai dan berbentuk gada. Tangkai anteridium dapat terdiri dari massa sel, misalnya A. laevis, atau dapat terdiri dari empat baris sel, seperti pada A. erectus dan A. punctatus. Antheridium yang tepat ditutupi oleh jaket berlapis tunggal. Di dalam jaket, terdapat banyak androsit yang akan bermetamorfosis menjadi antherozoid.
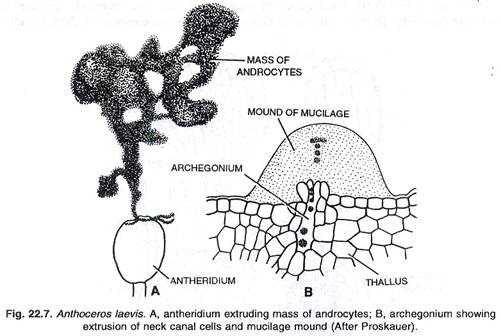
Pada pematangan antheridium, atap ruang antheridial hancur, akibatnya antheridia terekspos ke luar. Segera setelah itu, antheridia menyerap air dan meledak di ujung apikalnya, memberi jalan bagi antherozoid untuk bergerak keluar. Kadang-kadang androsit keluar dalam bentuk massa buram pada pembukaan anteridium. Dalam beberapa menit mereka bermetamorfosis menjadi antherozoids.
antherozoid:
Antherozoid berbentuk gelendong dan biciliate. Silia melekat pada ujung anterior tubuh. Kadang-kadang tepat di dekat titik perlekatan flagela ke tubuh, blepharoplast terlihat. Antherozoids berenang di air dengan momen cambuk dari flagela mereka.
Perkembangan arkegonium:
Archegonia ditemukan tertanam di thallus. Mereka tetap berhubungan langsung dengan sel vegetatif jaringan talus di sebelahnya. Mereka tidak memiliki sel steril jaket di sekitar mereka. Perkembangan archegonium dimulai dari satu sel superfisial. Sel ini menjadi menonjol dan bertindak sebagai inisial archegonial.
Menurut Mehra dan Handoo (1953), telah ditetapkan bahwa inisial archegonial berfungsi langsung sebagai sel archegonial primer. Dahulu diyakini (Campbell) bahwa ia membelah, dan menghasilkan dua sel, sel archegonial primer dan sel tangkai primer. Inisial archegonial pertama membelah secara vertikal, menghasilkan tiga inisial jaket yang mengelilingi sel aksial.
Sel aksial membelah secara melintang, menghasilkan inisial penutup dan sel pusat. Setelah itu, sel sentral membelah oleh dinding transversal, menghasilkan sel kanal primer dan sel venter primer. Sel kanal primer membelah berulang kali, menghasilkan file linier 4-6 sel kanal leher. Sel venter primer membelah sekali secara melintang, sehingga menimbulkan dua sel, sel kanal ventral dan telur (oosfer).
Relatif, sel saluran leher lebih sempit dari sel saluran venter dan sel telur. Akhirnya, tiga jaket inisial juga dibagi oleh dinding melintang. Perkembangan lebih lanjut dari lapisan jaket sama sekali tidak jelas. Karena sel-sel di permukaan bawah telur berasal dari inisial archegonial, mereka tidak dapat diperlakukan sebagai bagian dari archegonium.
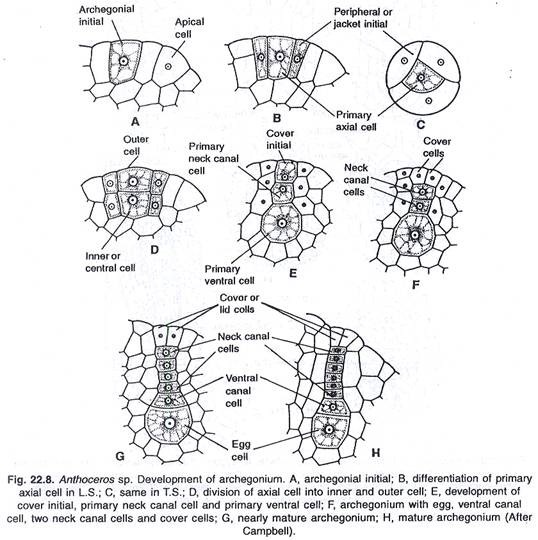
Struktur arkegonium:
Pada pematangan arkegonium, sel kanal venter dan sel kanal leher menjadi tergelatinisasi. Jadi arkegonium dewasa berbentuk seperti labu, tanpa sel saluran leher dan dengan telur (oosfer) di venternya. Di bagian atas leher arkegonium terdapat empat sel penutup, yang terpisah dari arkegonium, segera setelah gelatinisasi sel saluran venter dan leher selesai.
Pemupukan (singami):
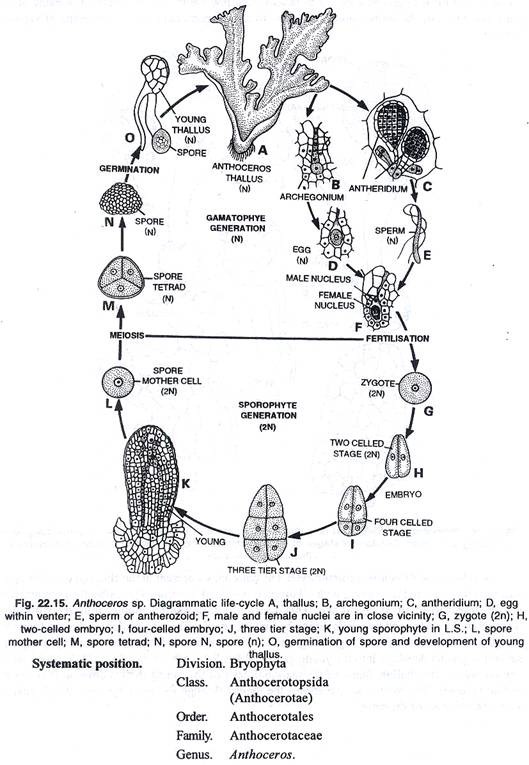
Sebelum pembuahan, sel-sel penutup terlepas dari arkegonium, dan sel-sel saluran leher menjadi gelatin. Melalui media air, antherozoids memasuki mulut arkegonium. Antherozoid tertarik secara kemotaktik. Pada akhirnya, satu antherozoid yang beruntung menembus sel telur, dan pembuahan dilakukan. Inti jantan dan betina bersatu satu sama lain, menghasilkan zigot (oospora), zigot adalah 2n, dan ini adalah awal dari tahap sporofit.
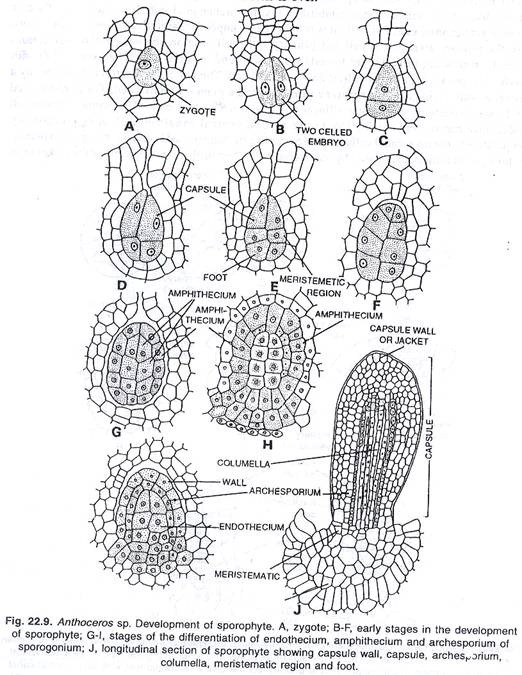
Perkembangan sporogonia:
Setelah pembuahan, zigot mengeluarkan dinding selulosa di sekitarnya dan membesar dalam ukuran yang masih mengisi venter arkegonium. Pembagian pertama zigot adalah vertikal. Namun menurut Pande (1932) dan Bhardwaj (1950), dalam kasus tertentu ini membelah secara melintang.
Dengan hasil pembelahan vertikal pertama, sel anak dihasilkan, yang mengalami pembelahan melintang menghasilkan empat sel dengan ukuran yang sama atau tidak sama. Ketika ukuran sel tidak sama, pasti sel menuju leher arkegonium adalah sel yang lebih besar.
Sel-sel ini kembali membelah secara vertikal, mengembangkan embrio bersel delapan, empat sel di setiap tingkat. Tingkat atas dari empat sel membelah secara melintang. Dengan cara ini masing-masing tiga tingkatan dari empat sel telah diproduksi. Tingkat paling bawah menghasilkan kaki, tingkat tengah menghasilkan sebagian kaki dan terutama seta dan tingkat paling atas menghasilkan kapsul.
Sel-sel pada tingkat paling bawah membelah secara teratur atau tidak teratur berkali-kali, menghasilkan kaki yang bulat. Kaki bersifat haustorial, yang menyerap makanan dari jaringan gametofit.
Tingkat paling atas dari empat sel, membelah satu atau dua kali secara melintang, menghasilkan dua atau tiga tingkat sel. Sekarang sel-sel membelah secara periklin, memunculkan lapisan luar amphithecium dan massa pusat sel endothecium. Pada sporogonium muda, columella terdiri dari empat baris vertikal sel, endothecium.
Pada sporogonium muda, columella terdiri dari empat baris vertikal sel, tetapi kemudian terdiri dari enam belas baris sel. Menurut Bhardwaj (1958) pada A. gemmulosus columella terdiri dari 36 sampai 49 baris vertikal sel. Amfitesium membelah secara periklin menghasilkan lapisan steril luar dari inisial jaket dan jaringan sporogen dalam, archesporium.
Inisial jaket membelah lagi dan lagi secara periklin menghasilkan dinding kapsul yang berlapis 4 sampai 6. Lapisan terluar berkembang menjadi epidermis berlapis tunggal. Sel-sel epidermis dipotong. Stomata mungkin atau mungkin tidak berkembang pada lapisan epidermis. Lapisan lainnya, di bawah epidermis berkembang menjadi klorenkim normal. Sel-sel klorofil ini, bagaimanapun, membantu mensintesis makanan.
Lapisan archesporium yang dikembangkan oleh divisi periclinal meliputi columella. Lapisan sporogen dapat memiliki ketebalan satu sampai empat sel dalam perkembangan selanjutnya. Pada Anthoceros hawaiensis ketebalannya tetap satu sel; pada A. pearsoni dan A. himalayensis dapat menjadi dua, tiga atau bahkan empat sel dengan ketebalan.
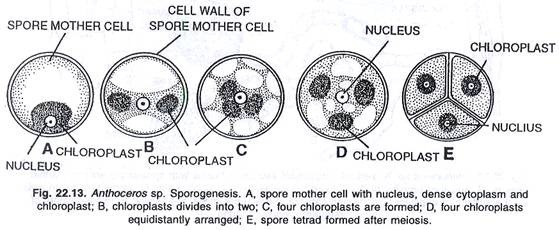
Pada tahap yang lebih muda, semua sel jaringan sporogen tetap berbentuk persegi panjang. Selanjutnya, jaringan sporogen berdiferensiasi menjadi dua jenis sel, yaitu (i) sporosit (sel induk spora) dan (ii) sel steril (pseudoelaters).
Sporosit atau sel induk spora mengalami pembelahan reduksi, masing-masing menghasilkan tetrad dari empat spora. Spora ini bersifat haploid. Spora, yang berkembang di bagian atas kapsul, matang terlebih dahulu. Setiap spora mengandung nukleus dan kloroplas.
Sel-sel steril segera membelah miring atau melintang menghasilkan tiga sampai lima pseudoelaters bersel. Saat jatuh tempo, pseudoelaters kehilangan protoplasma mereka. Dinding elater mungkin halus, menebal tidak teratur atau dengan penebalan spiral. Penebalan seperti itu, bervariasi dari spesies ke spesies. Pseudoelaters membantu dalam dehisensi spora dan berperilaku seperti elater sejati. Pada tahap awal fungsinya tampaknya bergizi.
Sporogonium dan dehisensinya:
Kapsul muncul dari thalli dalam bentuk struktur tanduk kecil. Biasanya panjangnya dua hingga tiga sentimeter. Tetapi pada beberapa spesies tingginya berkisar antara lima hingga lima belas sentimeter dan karena penampilan tanduknya, spesies ini disebut ‘lumut tanduk’. Sporogonium dewasa terdiri dari kaki bulat dan kapsul yang menonjol, ramping dan tegak. Ada zona meristematik di atas kaki, bukan seta.
Kaki bulat terdiri dari parenkim. Itu tetap menembus di thallus dan bertindak sebagai haustorium. Sel superfisial kaki seperti palisade. Ruang antara kaki dan kapsul ditempati oleh zona meristematik. Sel-sel di zona ini membelah sepanjang kecuali dan sampai makanan benar-benar habis. Dengan cara ini, kapsul bertambah panjang. Kapsul bertambah tinggi bahkan setelah matang.
Sel-sel yang berkembang dari meristem berdiferensiasi menjadi lapisan jaket, columella dan archesporium. Jadi, bagian atas sporogonium matang terlebih dahulu, dan bagian basal tetap muda. Kapsul tidak matang dengan kecepatan yang sama di semua bagiannya. Spora yang matang melepaskan diri dari bagian atas kapsul sedangkan pada bagian basal sel masih dalam keadaan embrionik.
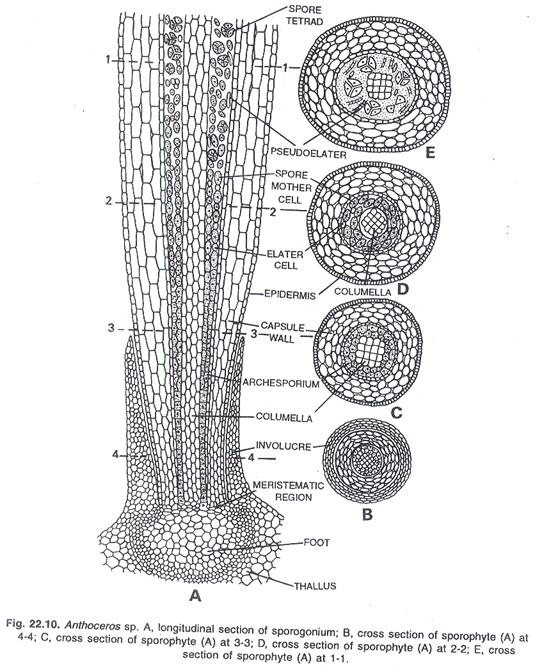
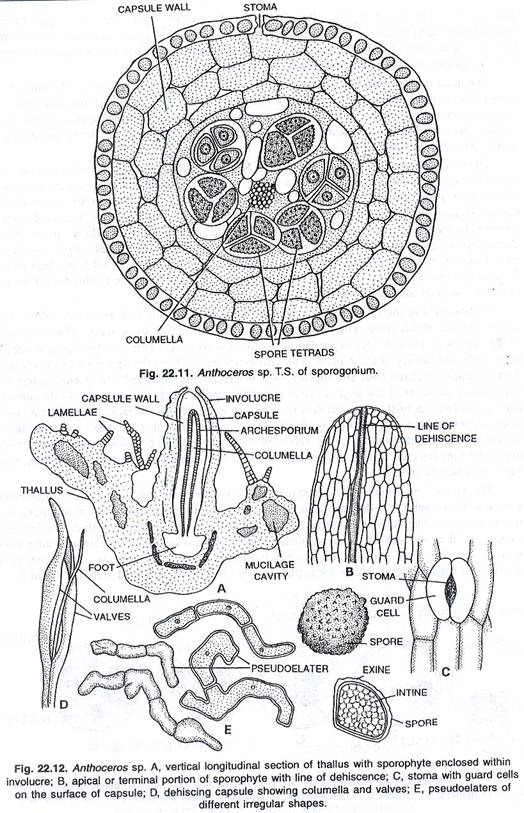
Kapsul utama terdiri dari banyak bagian penting. Wilayah tengah kapsul ditempati oleh columella steril. Menurut Campbell (1924), pada A. fusiformis columella berperan sebagai jaringan penghantar air. Namun, fungsi columella ini pada spesies lain tidak begitu pasti. Fungsi utama columella adalah memberikan dukungan mekanis pada sporogonium. Ini juga membantu dalam penyebaran spora.
Columella tetap dikelilingi oleh jaringan sporogenous. Di wilayah tepat di atas kaki, archesporium berlapis tunggal dan terlalu muda. Daerah ujung sporogonium memiliki spora dan elater yang matang. Dinding kapsul terdiri dari empat sampai enam lapisan sel parenkim. Lapisan terluar adalah epidermis, yang diselingi oleh stomata di beberapa tempat. Sel-sel epidermis dipotong.
Stomata terbuka di ruang antar sel sel klorofil. Biasanya setiap sel memiliki dua kloroplas. Proses fotosintesis berlangsung melalui kloroplas dan stomata. Dengan cara ini, makanan organik disintesis untuk sporogonium. Namun, sporogonium tetap bergantung pada thallus untuk suplai air dan nutrisi lainnya sepanjang hidupnya.
Saat matang, ujung sporogonium menjadi hitam atau coklat tua. Kapsul membelah pada tahap ini. Ujung kapsul mengerut karena kehilangan air. Dehiscence dari kapsul lebih atau kurang tergantung pada hilangnya air. Dengan cara ini suasana kering membantu dalam dehisensi kapsul.
Dehiscence dimulai dari daerah ujung kapsul. Mula-mula muncul celah memanjang kecil, yang melebar dan membesar. Pseudoelaters bersifat higroskopis. Mereka mulai berputar setelah eksposisi massa internal ke luar. Karena pelintiran pseudoelaters ini, tekanan diberikan pada lapisan selubung, dan ia meledak membebaskan spora di atmosfer.
Menurut spesies, dehiscence terjadi melalui satu hingga empat celah memanjang. Katup kapsul melengkung ke belakang dan akhirnya, saat mengering, mereka menjadi saling terpelintir. Spora yang dibebaskan disebarkan oleh angin dari satu tempat ke tempat lain.
Spora:
Pada tahap awal spora ditemukan tersusun dalam tetrad. Setelah dipisahkan satu sama lain, mereka bubar. Setiap spora agak bulat dan memiliki dua lapisan dinding. Lapisan dinding luar bersifat exine dan lapisan dinding dalam bersifat intine.
Bagian dalamnya halus dan tipis, sedangkan bagian luarnya agak tebal dan berornamen. Warna spora dewasa bervariasi dari satu spesies ke spesies lainnya; ini mungkin kuning, coklat, coklat tua atau hitam. Setiap spora memiliki inti tunggal, plastida tidak berwarna, sedikit tetesan minyak dan bahan makanan di dalamnya.
Perkecambahan spora:
Setelah dibebaskan dari sporogonium, spora mengalami masa istirahat sebelum berkecambah yang berkisar dari beberapa minggu hingga beberapa bulan. Bagian luar spora pecah, dan bagian dalam keluar dalam bentuk tabung germinal atau protonema dengan panjang bervariasi.
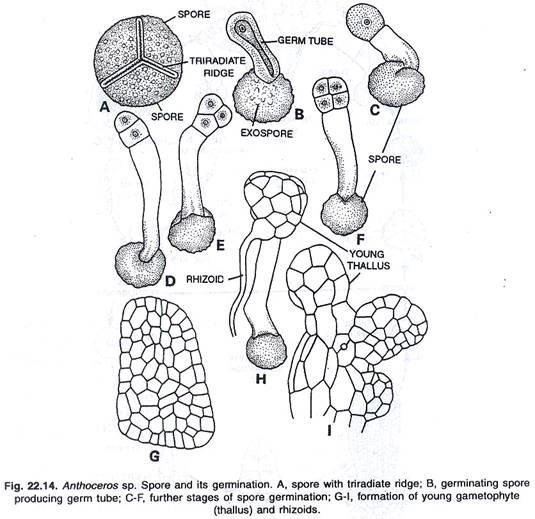
Perkembangan gametofit muda:
Klorofil yang ada dalam kloroplas spora melewati tabung germinal bersama dengan tetesan minyak dan bahan makanan. Setelah itu tabung germinal membelah secara melintang di ujung apikalnya. Pembagian ini diikuti oleh pembagian melintang lainnya. Segera setelah itu, dua sel yang terbentuk membelah oleh dinding yang berpotongan memanjang dan kuadran dihasilkan.
Di ujung terminal muncul titik tumbuh dengan sel apikal yang dengan menghasilkan beberapa segmen berkembang menjadi gametofit muda. Segera setelah celah lendir berkembang di sisi ventral thallus. Beberapa sel marjinal talus muda berkembang menjadi rizoid berdinding halus. Nostoc sp. menembus talus melalui celah lendir, yang kemudian membentuk koloni yang sama.
Hubungan antar Anthocerotopsida:
Anthocerotopsida berbeda dari bryophyta lain dalam beberapa hal. Perbedaan tersebut adalah:
(a) Di antara Anthocerotopsida sel-selnya memiliki kloroplas besar dan setiap kloroplas mengandung pirenoid.
(b) Pada kelompok ini perkembangan anteridia terjadi dari sel-sel hipodermis pada sisi dorsal gametofit.
(c) Archegonia kelompok ini hampir seluruhnya tertanam dalam gametofit.
(d) Pertumbuhan sporofit tidak dapat ditentukan karena daerah meristematik terus bertambah pada dasar kapsul.
Karena perbedaan yang disebutkan di atas, kelas ditempatkan di antara Hepaticopsida di satu sisi dan Bryopsida di sisi lain.
Fitur lain dari minat khusus Anthocerotopsida adalah:
(a) Gametofit berbentuk thallose, agak bercuping atau membedah secara radial, dan terkadang menunjukkan kecenderungan percabangan dikotomis.
(b) Gametofit selalu berdiferensiasi dorsiventral dan memiliki banyak rizoid berdinding halus pada permukaan ventral.
(c) Permukaan ventral tidak memiliki sisik dan rambut lendir.
(d) Margin lateral sebagian besar genera kelompok ini (kecuali Dendroceros) memiliki ketebalan lebih dari satu sel.
(e) Tidak ada diferensiasi internal jaringan.
(f) Bagian ventral thallus memiliki rongga antar sel yang berisi lendir di sisi dorsal dan membuka ke permukaan melalui celah sempit. Rongga ini umumnya mengandung koloni Nostoc (ganggang hijau biru) di dalamnya.
(g) Pirenoid dari Anthocerotopsida tidak homolog dengan alga hijau (chlorophyceae) karena terdiri dari massa yang padat dari 25 hingga 300 cakram atau badan berbentuk gelendong (McAllister, 1914, 1927).
(h) Pertumbuhan thallus diawali oleh satu sel apikal dengan dua permukaan pemotongan (kecuali pada Dendroceros di mana terdapat tiga permukaan pemotongan).
(i) Reproduksi vegetatif melalui kematian dan pembusukan thalli lebih jarang terjadi pada kelompok ini dibandingkan dengan Hepaticopsida. Namun, pembentukan umbi sering terjadi pada kelompok ini.
(j) Sebagian besar spesies homothallic, tetapi beberapa heterothallic (Proskauer, 1948). Pada spesies heterothallic, penentuan jenis kelamin bersifat genotipik, yaitu dua spora tetrad berkembang menjadi jantan dan dua menjadi gametofit betina.
(k) Anthocerotopsida dan Hepaticopsida pada dasarnya berbeda dalam fakta bahwa pada yang pertama inisial antheridial adalah sel anak dalam yang dihasilkan oleh pembelahan periclinal dari sel anak superfisial gametofit. Hal ini menunjukkan bahwa Anthocerotopsida berasal dari nenek moyang dimana antheridia berkembang dari sel dorsal superfisial.
(l) Dalam Anthoceros dan beberapa genera lain dari kelompok ini, inisial antheridial dapat membelah secara vertikal menjadi dua atau empat sel anak, yang masing-masing berkembang menjadi antheridium.
(m) Perkembangan sel antheridial primer menjadi antheridium yang sebenarnya sama seperti di Sphaerocarpales dan Marchantiales.
(n) Pada Anthocerotopsida, spermatogenesis sangat mirip dengan bryophyta lain dan melibatkan metamorfosis androsit menjadi sperma biflagellata pada antherozoid.
(o) Pada Anthocerotopsida fungsi awal archegonial langsung sebagai sel archegonial primer alih-alih membelah menjadi sel archegonial primer dan sel tangkai primer seperti yang ditemukan pada lumut lainnya.
(p) Pembelahan pertama zigot adalah vertikal tetapi kasus telah ditemukan (Bhardwaj, 1950; Pande, 1932) di mana pembelahan melintang terjadi.
(q) Amfitesium membelah secara periklin, dimana lapisan luar berfungsi sebagai lapisan awal selubung dan lapisan dalam sebagai arkesporium.
(r) Salah satu ciri unik Anthocerotopsida adalah bahwa sel-sel kapsul tidak matang pada kecepatan yang sama dan sel-sel di bagian basal kapsul tetap menjadi embrionik bahkan setelah sel-sel di bagian apikal matang sepenuhnya. Fitur ini tidak ditemukan pada bryophyta lain.
Afinitas Anthocerotopsida:
Kelompok ini menunjukkan kesamaan dengan ganggang hijau, Hepaticopsida (lumut hati), Bryopsida (lumut) dan Psilophytales dari Pteridophyta.
(A) Fitur umum dengan ganggang hijau:
Ganggang hijau (Chlorophyceae) telah dianggap sebagai kelompok asal bryophyta dan pteridophyta.
(i) Biasanya setiap sel gametofit memiliki satu kloroplas besar dengan bentuk tertentu.
(ii) Pirenoid hadir dalam kloroplas sel gametofit. Kehadiran pirenoid adalah karakteristik sel ganggang hijau saja
(iii) Pirenoid dari Anthocerotopsida dan alga hijau memiliki fungsi yang sama dan membentuk butiran pati di daerah periferalnya.
(iv) Garis besar dan percabangan gametofit serupa dalam kedua kasus.
(v) Adanya antherozoid biciliate atau biflagellate (keduanya flagella tipe whiplash).
(B) Fitur umum dengan lumut hati (Hepaticopsida):
Banyak ahli taksonomi memasukkan Anthocerotales di Hepaticae.
(i) Gametofitnya seperti thallus.
(ii) Rizoid berdinding halus ditemukan pada anggota Anthocerotopsida serta anggota Jungermanniales dari Hepaticopsida.
(iii) Pertumbuhan apikal thallus serupa pada keduanya.
(iv) Archesporium memunculkan spora dan sel steril pada kedua kelompok. Sel-sel steril dengan pita spiral ditemukan di Megaceros dari Anthocerotopsida yang menunjukkan kesamaan dengan banyak anggota Hepaticopsida.
(v) Diferensiasi amphithecium dan endothecium oleh dinding periclinal mirip dengan banyak Hepaticae.
(C) Fitur umum dengan Bryopsida (lumut):
(i) Adanya kolumela sentral pada kedua kelompok.
(ii) Pengurangan besar jaringan sporogen pada kelompok Anthocerotopsida dan Bryopsida.
(iii) Kehadiran stomata fungsional (misalnya, di Funaria).
(iv) Diferensiasi archesporium dari amphithecium bagian dalam seperti pada Sphagnales. Fitur ini menunjukkan hubungan antara Anthocerotopsida dan Bryopsida.
(v) Tahap perkembangan embrio sangat mirip. Pembagian awal sangat mirip.
(D) Fitur umum dengan Pteridophyta:
(i) Adanya organ seks yang cekung umum terjadi pada kedua kelompok.
(ii) Adanya struktur vegetatif yang serupa dari gametofit di Anthoceros dan Fern.
(iii) Adanya sporogonium yang sangat berkembang dengan pertumbuhan tak tentu jaringan fotosintetik dan stomata fungsional.
Fakta-fakta tersebut di atas mendukung bahwa Anthocerotopsida adalah kelompok tanaman yang berbeda tetapi sintetik. Ini membentuk tautan penghubung dengan lumut hati dan lumut di satu sisi dan dengan pteridofit di sisi lain. Ada juga koneksi jarak jauh dengan ganggang hijau (Chlorophyceae). Campbell (1928) menyarankan, “Fakta bahwa jaringan sporogenous primer di Anthocerotales selalu muncul dari amphithecium, sementara di semua lumut hati lainnya dikembangkan dari endothecium, tampaknya menjadi perbedaan yang radikal”.
Campbell juga berpendapat bahwa sporofit Anthoceros dengan sistem asimilasinya dengan stomata dan pertumbuhan yang berkelanjutan menunjukkan aliansi yang erat dengan sporofit bercabang dikotomis yang independen dan tidak berakar dari kelompok fosil primitif, Psilophytales. Mehra (1957) menyatakan bahwa Anthocerotopsida dan Psilopsida muncul dari stok Anthorhyniaceae biasa.
Signifikansi biologis Anthocerotopsida:
Sporofit Anthocerotopsida menunjukkan kemungkinan garis kemajuan biologis sebagai berikut:
(i) Kehadiran sistem asimilasi berventilasi yang rumit menunjukkan awal dari kemandirian fisiologis sporofit.
(ii) Penghentian jaringan sporogen terus menerus oleh pertumbuhan sel steril antara sel induk spora yang menunjukkan awal pembentukan sporangia.
(iii) Pembentukan kolumela steril yang berkembang dengan baik dari endotelium sentral menunjukkan awal pembentukan sistem konduksi, dan tahap awal pembentukan sporangia superfisial.
(iv) Kehadiran zona meristematik interkalar menunjukkan awal dari pertumbuhan sporofit yang tidak pasti dan terus menerus.
Ringkasan:
Posisi dan distribusi sistematis:
Divisi-Bryophyta; kelas Anthocerotopsida (Anthocerotae); Orde-Anthocerotales; Keluarga- Anthocerotaceae; Genus-Anthoceros. 25 spesies ditemukan di India; kosmopolitan dalam distribusi.
Kebiasaan: Fitur eksternal:
Tempat teduh yang lembab. Thallus tipis, dorsiventral dengan tulang rusuk tengah yang tidak jelas; kecenderungan percabangan dikotomis; rizoid-uniseluler, hanya berdinding halus; sisik tidak bercabang tidak ada.
Struktur internal:
Semua sel berfotosintesis; pertumbuhan apikal baik oleh satu sel apikal atau sekelompok sel apikal.
Organ seksual:
Berumah satu dan dioecious; antheridia endogen, di dalam rongga antheridial tertutup pada permukaan dorsal thallus, tangkai panjang, ramping atau masif, selubung tubuh berlapis tunggal, antherozoid biciliate; archegonia-tertanam di permukaan dorsal thallus, leher terdiri dari 6 baris vertikal sel, venter setebal satu lapis, sel penutup 2 sampai 4, sel saluran leher biasanya 4.
Sporofit:
Amphithecium memunculkan selubung kapsul dan archesporium; endotelium memunculkan columella; archesproium menimbulkan sporosit dan pseudoelaters; kaki di sini bulat; seta tidak ada; kapsul panjang dan silindris dan dindingnya berlapis empat sampai enam dan kokoh; pseudoelaters tanpa pengental; columella hadir, melengkung oleh kantung spora; Dehiscence spora terjadi ketika dinding kapsul terbagi menjadi 1 sampai 4 katup, yang tetap bersatu di ujungnya, pseudoelaters membantu penyebaran spora.
Gametofit muda:
Dua lapisan dinding spora; ukuran spora 0,025-0,05 mm; tabung germinal terbentuk pada saat spora berkecambah.



