Persilangan Kromosom: Mekanisme, Macam, Faktor dan Signifikansi!
Crossing over adalah proses pertukaran materi genetik atau segmen antara kromatid non-saudara dari dua kromosom homolog. Persilangan terjadi karena pertukaran bagian kromosom homolog.
Biasanya, jika pemilahan independen terjadi yaitu ketika gen hadir pada kromosom yang berbeda, kita harus mengharapkan rasio uji silang 1:1:1:1. Tetapi ketika kita melihat Gambar 5.48, jumlah kelas rekombinan yang muncul sangat sedikit. Oleh karena itu dapat disimpulkan bahwa dua gen berada pada kromosom yang sama dan munculnya rekombinan dalam jumlah rendah dihasilkan dari persilangan.
Kromosom biasanya mengalami kerusakan selama gametogenesis. Jadi, memang ada mekanisme di mana sekelompok gen pada kromosom yang sama berubah dengan kelompok gen serupa pada kromosom homolog. Persentase pindah silang yang diperoleh antara gen-gen terpaut berbeda bervariasi menurut jarak antar gen pada kromosom.
Lebih lanjut, kedua gen terpisah pada kromosom, lebih mungkin terjadi persilangan di antara keduanya. Pada titik tertentu hanya dua dari empat kromatid yang terlibat dalam pertukaran bagian mereka dan menghasilkan 50 persen gamet rekombinan.
Gen-gen dalam kasus seperti itu harus terletak jauh terpisah pada kromosom yang sama untuk memungkinkan persilangan di semua sel induk selama pembelahan reduksi. Dalam kondisi seperti itu, gen berperilaku seolah-olah terletak pada kromosom yang berbeda. Hukum Mendel Assortment Independen berlaku hanya di bawah kondisi berikut:
(a) Ketika gen terletak pada kromosom yang berbeda.
(b) Jika gen-gen tersebut terletak pada kromosom yang sama tetapi jarak antara keduanya sangat baik sehingga menghasilkan 50 persen gamet rekombinan akibat persilangan.
Mekanisme penyeberangan:
Mekanisme pindah silang dapat dijelaskan di bawah judul berikut:
(i) Sinapsis:
Kromosom homolog berpasangan memanjang karena gaya tarik-menarik timbal balik dalam zigot profase-I pada meiosis. Pemasangan dimulai pada satu titik atau lebih dan berlanjut sepanjang panjang dengan gaya ritsleting. Proses berpasangan disebut sinapsis. Kromosom homolog berpasangan disebut bivalen. Selama sinapsis, perancah molekuler yang disebut kompleks sinaptonemal menyelaraskan molekul DNA dari dua kromosom homolog berdampingan.
(ii) Duplikasi kromosom:
Sinapsis diikuti oleh duplikasi kromosom yang mengubah sifat bivalen kromosom menjadi tahap empat beruntai atau tetravalen. Tahap empat untai (Gambar 5.48) kromatid terjadi karena pemisahan kromosom homolog menjadi kromatid saudara yang terikat dengan sentromer yang tidak terbelah.
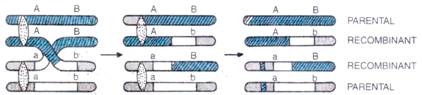
Gambar 5.48. Persilangan pada tahap untai 4 menghasilkan 50% rekombinan dan 50% tipe gamet parental.
(iii) Penyeberangan:
Pada pachytene, persilangan terjadi. Kromatid non-saudara dari pasangan homolog saling melilit karena aksi enzim endonuklease. Kromatid terhubung satu sama lain pada titik yang dikenal sebagai chiasmata.
Penyeberangan bisa dilakukan di beberapa titik. Jumlah chiasmata yang terbentuk sebanding dengan panjang kromatid. Gen-gen pada lokus yang jauh mengalami persilangan tetapi gen-gen yang ditempatkan dekat gagal melakukan persilangan dan menunjukkan fenomena pertalian.
Selama diakinesis profase-I chiasmata bergerak menuju akhir bivalen dengan proses yang disebut terminalisasi. Dengan demikian kromatid puntir terpisah sehingga kromosom homolog terpisah sepenuhnya.
Pada anafase – 1 meiosis, kromosom homolog terpisah. Jelaslah bahwa salah satu kromatid dari setiap kromosom membawa sebagian kromatid dari kromosom homolognya. Pada akhir meiosis, empat jenis gamet terbentuk. Dua akan menjadi tipe induk dan dua akan berisi kromosom dengan rekombinasi gen yang terbentuk selama persilangan.
Janssens (1909) adalah orang pertama yang memahami dengan benar proses pembentukan kiasma. Apa yang sebenarnya menyebabkan kerusakan dan penyatuan kembali kromatid masih belum jelas. Menurut Stem dan Hota (1978) pecah atau torehan muncul pada kromatid akibat enzim endonuklease.
Torehan ini biasanya tersumbat tetapi satu dari 1000 mengembangkan celah dengan bantuan enzim eksonuklease. Segmen kromatid terpisah di antara dua celah dengan bantuan enzim yang disebut U-protein. Segmen-segmen ini bersatu kembali dengan bantuan R-protein.
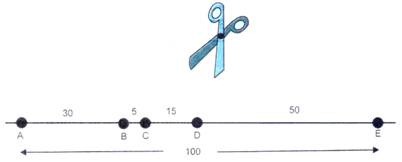
Gambar 5.49. Lima simpul A, B, C, D dan E pada seutas tali dipisahkan oleh jarak yang ditunjukkan.
Crossing-over dan chiasmata:
Ada dua teori tentang hubungan antara persilangan dan pembentukan chiasmata.
- Teori tipe chiasma (Jenssen, 1909):
Menurut teori ini, tindakan pindah silang diikuti oleh pembentukan chiasma. Di sini formasi kiasma adalah konsekuensi dari persilangan. Pandangan ini menyatakan bahwa loop yang berdekatan diatur dalam satu bidang dan karenanya disebut teori satu bidang. Menurut teori ini, pindah silang terjadi pada tahap pakiten dan kiasma muncul pada tahap diploten.
- Teori klasik (Sharp, 1934):
Menurut teori ini, persilangan merupakan hasil pembentukan kiasma. Loop yang berdekatan diatur pada sudut kanan satu sama lain dan karenanya disebut teori dua bidang. Chiasmata diatur pada pachytene dan pindah silang terjadi pada tahap diplotene. Teori ini telah dianggap tidak dapat dipertahankan dan karenanya ditolak.
Jenis Crossing-over:
Bergantung pada jumlah chiasmata yang muncul, jenis persilangan berikut dapat didiskusikan:
(i) Penyeberangan tunggal:
Dalam hal ini, hanya satu chiasma yang terbentuk yang mengarah pada pembentukan gamet persilangan tunggal. Ini adalah jenis cross-over yang paling umum.
(ii) Persilangan ganda:
Dalam persilangan ganda, dua chiasmata berkembang. Chiasmata ini dapat muncul di antara kromatid yang sama atau di antara kromatid yang berbeda. Jenis persilangan ini membentuk gamet persilangan ganda.
(iii) Persilangan ganda:
Di sini, ‘terbentuk lebih dari dua chiasmata. Ini dapat diklasifikasikan lebih lanjut menjadi rangkap tiga (3 chiasmata), quadruple (4 chiasmata) dan seterusnya. Persilangan ganda jarang terjadi.
Faktor-faktor yang mempengaruhi pindah silang:
Jarak antar gen. Semakin jauh jarak antara dua gen pada kromosom yang sama, semakin tinggi frekuensi pindah silang.
Signifikansi penyeberangan:
(i) Proses ini menyediakan simpanan variabilitas genetik yang tidak habis-habisnya dalam organisme yang bereproduksi secara seksual.
(ii) Rekombinasi yang berguna digunakan oleh pemulia tanaman dan hewan. Peternak mencoba memutuskan pertalian dengan menyilangkan untuk mendapatkan kombinasi sifat-sifat yang berguna dalam keturunannya.
(iii) Proses ini menghasilkan kombinasi gen baru (rekombinasi). Revolusi hijau dan revolusi putih terutama disebabkan pengambilan selektif dari rekombinasi genetik yang berguna yang dikembangkan oleh proses persilangan.



