Berikut adalah catatan Anda tentang Kromosom Prokariotik!
Informasi genetik sel prokariotik dibawa dalam nukleoplasma pada struktur yang disebut kromosom bakteri. Ini terdiri dari molekul DNA heliks ganda (asam deoksiribonukleat), tidak pernah dikaitkan dengan protein dasar, dan telah ditunjukkan pada beberapa prokariota berbentuk lingkaran.
Gambar Courtesy: hangthebankers.com/wp-content/chromosome.jpg
Kromosom bakteri akibatnya tidak homolog struktural dengan kromosom nuklir dari sel eukariotik, melainkan dengan DNA organel hadir dalam mitokondria eukariotik dan kloroplas. Ada kemungkinan bahwa satu kromosom bakteri (yaitu, satu molekul DNA yang sangat panjang) membawa semua informasi genetik yang diperlukan untuk menentukan sifat-sifat esensial sel prokariotik.
Plasmid: 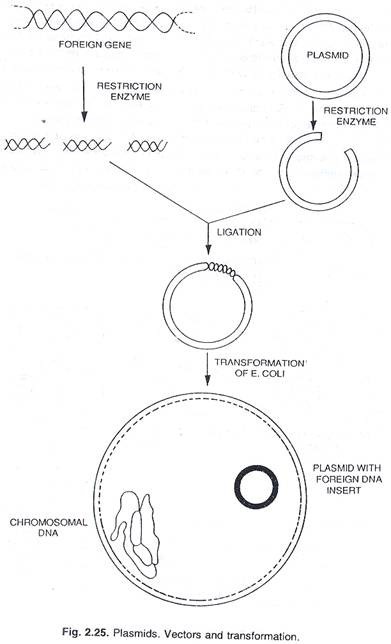
Banyak bakteri juga dapat menampung molekul DNA sirkuler ekstra kromosom kecil yang mampu melakukan replikasi otonom, yang dikenal sebagai plasmid yang sejauh ini diselidiki membawa determinan untuk karakter fenotipik seperti resistensi terhadap obat dan zat antibakteri lainnya, dan untuk enzim yang memediasi jalur metabolisme tertentu. . Jumlah DNA dalam plasmid adalah dari 0,1 sampai 5 persen dari yang ada dalam kromosom bakteri.
Ini ditemukan di Escherichia coli bahwa ada dua jenis perkawinan, dan selama konjugasi satu pasangan hanya bertindak sebagai donor genetik, atau laki-laki, dan yang lainnya hanya sebagai penerima genetik atau perempuan. Karena satu-satunya fungsi jantan adalah untuk mentransfer sebagian DNA-nya, sel itu tidak perlu tetap hidup, sedangkan sel betina harus tetap hidup, sehingga zigot dapat berkembang di dalamnya.
Ditemukan bahwa kejantanan pada bakteri ditentukan oleh unsur genetik yang dapat ditularkan ketika bakteri jantan dan betina berkonjugasi, setiap sel betina diubah menjadi jantan. Elemen genetik yang mengatur sifat kelelakian yang diwariskan disebut faktor F (F = fertilitas), yang ditransmisikan hanya melalui kontak sel ke sel secara langsung. Pada tahun 1952 Lederberg menciptakan istilah plasmid sebagai nama genetik untuk semua determinan herediter ekstra kromosom, di mana F adalah contohnya.
Sekarang diketahui bahwa plasmid bakteri adalah molekul kecil melingkar dari DNA yang membawa gen untuk replikasinya sendiri. Dalam banyak kasus, mereka juga membawa gen yang memberikan sifat baru pada sel inang, seperti resistensi terhadap obat atau produksi racun. Banyak plasmid membawa gen yang mengatur proses konjugasi. Jadi, konjugasi adalah mekanisme yang dikenakan pada sel bakteri oleh plasmid, hasil normalnya adalah transfer DNA plasmid.
Jenis plasmid:
Sebagian besar plasmid telah diklasifikasikan berdasarkan sifat inangnya. Jadi, ada faktor R (R = resistensi) dan faktor COI (COI = colicinogeny) dari bakteri gram negatif, plasmid penisilinase Staphylococcus aureus, plasmid degradatif Pseudomonas, plasmid kriptik, dll.
Properti plasmid:
Struktur semua plasmid yang diketahui terdiri dari molekul DNA beruntai ganda yang melingkar. Beberapa dari mereka memiliki berat molekul dalam kisaran 5 × I0 7 sampai 7 × 10 7 . Satu (faktor R) memiliki berat molekul hanya 1 × 10 7 , dan beberapa plasmid samar bahkan lebih kecil. Karena jumlah DNA yang dibutuhkan untuk mengkode rata-rata polipeptida dengan berat molekul 40.000 adalah sekitar 6 × 10 7 , F 1 dan plasmid lain dengan ukuran yang sama dapat mengandung sebanyak 100 gen.
Ekspresi Gen pada Prokariota:
Kromosom bakteri:
Telah ditetapkan sekarang bahwa kromosom bakteri selalu ada di tengah sel bakteri yang tidak mampu membelah mitosis dan meiosis dan tidak tertutup dalam membran inti tertentu. Sel bakteri rata-rata mengandung seperseribu isi DNA dari sel eukariotik yang khas.
Kromosom bakteri adalah filamen molekul DNA tipis memanjang fleksibel dan melingkar yang terkait dengan beberapa protein. Sel bakteri dapat membelah dengan sangat cepat. Pembelahan sel diselesaikan dengan menggandakan semua konstituen sel diikuti dengan partisi sel menjadi dua sel anak. Langkah pertama dalam pembelahan sel adalah duplikasi DNA.
Rekombinasi Genetik pada Bakteri:
Ini adalah proses di mana materi genetik, yang terkandung dalam dua genom terpisah, disatukan dalam satu unit. Pada bakteri rekombinasi terjadi dengan (1) transformasi, (2) transduksi dan (3) konjugasi. Fenomena ini telah dibahas secara rinci dalam paragraf sebelumnya, di bawah judul ‘reproduksi seksual pada bakteri’.
Induksi dan Represi:
Potensi genetik virus terbatas dan pada dasarnya menggunakan mesin inang untuk replikasinya. Di sisi lain, bakteri dapat mensintesis konstituennya dari garam dan gula sederhana. Bakteri, Escherichia coli memiliki sekitar 2500 gen pada kromosomnya. Bakteri mempertahankan ekonomi internalnya hanya dengan membiarkan aksi beberapa gen pada satu waktu.
Ini berarti bahwa gen akan dihidupkan atau dimatikan sesuai kebutuhan. Satu set gen akan diaktifkan ketika ada kebutuhan untuk menangani dan memetabolisme substrat baru. Ketika set gen ini diaktifkan, enzim diproduksi, yang memetabolisme substrat baru. Fenomena ini disebut induksi dan molekul (metabolit) yang menimbulkan fenomena ini adalah penginduksi.
Di sisi lain, ketika metabolit yang dibutuhkan oleh bakteri disuplai secara berlebihan dari luar, bakteri menghambat pembuatannya dan ini menghentikan produksi metabolit lebih lanjut oleh bakteri. Gen-gen yang tidak aktif ini dinamai represibel, dan fenomena ini dikenal sebagai represi atau represi umpan balik.
Namun, beberapa gen secara konstan diekspresikan untuk menjaga aktivitas seluler normal seperti glikolisis. Gen-gen ini dikenal sebagai konstitutif, dan enzim yang dihasilkannya dikenal sebagai enzim konstitutif, seperti dehidrogenase.
Konsep Operon:
Francois Jacob dan Jacques Monod (1961), saat bekerja di Institut Pasteur, Paris, mengusulkan bahwa jalur metabolisme diatur sebagai satu kesatuan. Mereka mempelajari perubahan sifat enzim E. coli ketika diberi berbagai jenis gula. Mereka menemukan, ketika laktosa gula ditambahkan ke kultur E. coli, itu menginduksi tiga enzim yang diperlukan untuk memecah laktosa menjadi glukosa dan galaktosa.
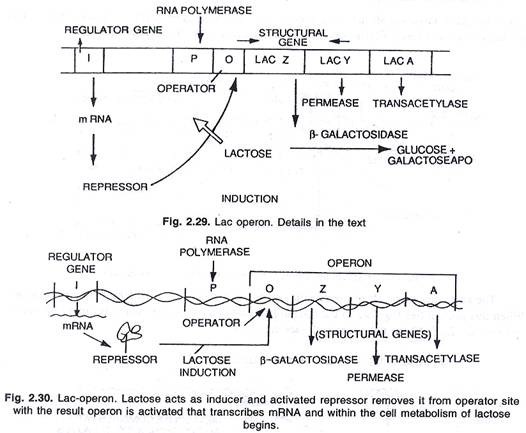
Enzim yang baru disintesis ini adalah- (i) p-galaktosidase, (ii) permease dan (iii) transasetilase. Menurut mereka, sintesis ketiga enzim ini dikendalikan oleh segmen DNA panjang yang dikenal sebagai operon yang dapat dibagi menjadi situs operator O dan tiga gen struktural Z, Y dan A. Aksi gen struktural diatur oleh situs operator dengan bantuan protein represor yang dihasilkan oleh aksi gen ‘i’ yang dikenal sebagai gen pengatur.
Gen diekspresikan atau tidak diekspresikan yang tergantung pada apakah sakelar operator hidup atau mati. Saat sakelar hidup, ketiga gen tersebut ditranskripsi oleh RNA polimerase menjadi rangkaian tunggal messenger RNA yang mencakup ketiga gen tersebut. Setiap gen struktural umumnya dikenal sebagai cistron dan mRNA panjang yang ditranskripsi yang menutupi semua cistron dikenal sebagai polikistronik.
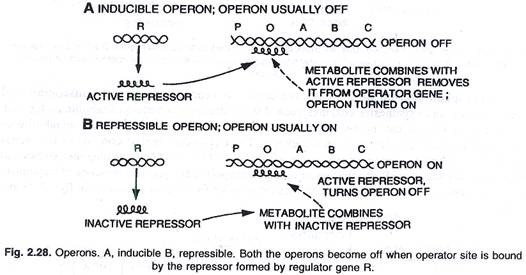
Pengaktifan atau penonaktifan sakelar operator dicapai oleh protein yang dikenal sebagai represor. Ketika protein ini berikatan dengan operator (O) dan memblokirnya, sakelar dimatikan dan ketiga gen (Z, Y, A) tidak diekspresikan, (lihat gbr. 2.28).
Lac-operon:
Ini adalah contoh operon yang dapat diinduksi. Beberapa molekul laktosa ditambahkan ke dalam sel oleh aksi enzim permease, jumlah kecil yang ditemukan bahkan dalam kondisi tertekan. Beberapa molekul ini kemudian diubah menjadi bentuk aktif laktosa yang berikatan dengan represor (produk gen i) dan dengan demikian, represor tidak dapat lagi berikatan dengan sakelar operator.
Ketika operator bebas dari represor RNA polimerase, ia memulai transkripsi operon dengan mengikat situs promotor P. Messenger RNA yang sesuai dengan ketiga enzim disintesis yang diterjemahkan untuk menghasilkan tiga enzim-P-galactosidase, permease dan transacetylase.
Dengan produksi ketiga enzim ini, metabolisme laktosa dimulai. Sintesis enzim dilanjutkan kecuali dan sampai semua molekul laktosa dikonsumsi oleh sel. Ketika molekul terakhir laktosa, yang terikat pada represor juga dikonsumsi, represor yang tidak aktif menjadi aktif dan berikatan dengan situs operator (O) untuk mematikan operon seperti biasa, (lihat gambar 2.29 dan 2.30).
Triptofan-operon:
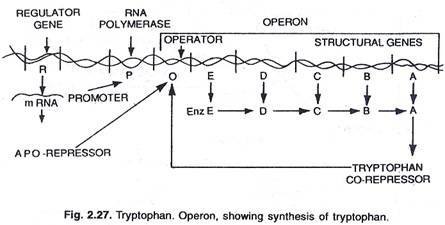
Dalam hal ini, operon umumnya aktif sehingga transkripsi dan translasi aktif untuk mensintesis enzim yang diperlukan untuk sintesis metabolit tertentu oleh sel. Namun, operon dapat dimatikan ketika sel tidak membutuhkan metabolit atau metabolit telah diproduksi secara berlebihan. Operon triptofan terdiri dari lima gen (trp E, D, C, B dan A) yang mengkode lima enzim (En z E, D, C, B, dan A) mengkatalisis sintesis triptofan (asam amino) dan dengan demikian membentuk jalur anabolik. Kehadiran triptofan berfungsi untuk menekan sintesis enzim yang bertanggung jawab untuk pembuatannya, (lihat gambar 2.27 dan 2.31).
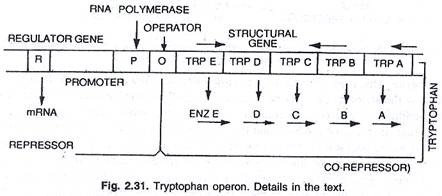
Lima gen struktural yang mengkode kelima enzim ini diatur oleh satu tombol operator. Di sini, protein represor yang diproduksi oleh gen pengatur tidak mengikat operator dengan sendirinya. Namun, ketika triptofan ditambahkan, ia berikatan dengan represor dan dikenal sebagai korepresor. Kompleks represor-korepresor berikatan dengan operator dan memblokir ekspresi operon ini.
Struktur operon triptofan kurang lebih mirip dengan operon lac, tetapi memiliki variasi fungsional. Dalam hal ini produk gen R setara dengan gen i dari lac operon. Produk gen R menghasilkan protein yang dengan sendirinya tidak dapat berikatan dengan operator. Ini disebut sebagai apo-represor.
Dengan adanya triptofan sebagai ko-represor, represor fungsional terbentuk yang sekarang berikatan dengan operator dan mencegah transkripsi operon dan produksi triptofan. Di sini model operon menjelaskan fenomena induksi dan represi pada prokariota.



