Baca artikel ini untuk mendapatkan informasi tentang kompetensi, determinasi, dan diferensiasi jaringan!
Kompetensi:
Waddington (1932) memperkenalkan konsep kompetensi sebagai keadaan fisiologis jaringan, yang memungkinkannya bereaksi dengan cara spesifik morfogenetik terhadap rangsangan determinatif. Itu selalu terkait dengan rangsangan tertentu dan tanggapan tertentu yang sesuai. Kompetensi adalah istilah yang merangkum kemampuan pelengkap enzim sel embrionik untuk mengadopsi rasio metabolit tertentu.
Gambar milik: mattek.com/uploaded/EpiDermFT/EpiDermFT%20Final/EpiDerm-FT2%20label.png
Ketika ektoderm embrio amfibi ditransplantasikan dari berbagai tahap perkembangan blastula ke neurula awal, secara bertahap kehilangan kompetensi saraf. Dengan penuaan, ektoderm secara bertahap kehilangan kemampuannya untuk merespons rangsangan induktif chordamesoderm.
Oleh karena itu, menurut Needham (1942) sel-sel pada tahap embrionik awal mungkin refrakter terhadap rangsangan induktif dan mencapai tahap inertness relatif terhadap induksi embrio spesifik tertentu pada tahap selanjutnya, yang masing-masing merupakan pra-kompetensi dan pasca-kompetensi (Lovtrup 1947). . Ektoderm terisolasi yang tidak terpajan pada induksi saraf, dan ektoderm yang ditransplantasikan terlambat, berdiferensiasi menjadi epidermis saja (Gbr. 1).
Dalam sebuah percobaan, penurunan kompetensi saraf dengan penuaan jaringan telah diuji dengan isolasi untuk berbagai jangka waktu dan mentransplantasikannya ke lokasi yang berbeda di neurula. Diamati bahwa kompetensi untuk membentuk struktur otak menurun pada tahap gastrula akhir dan ektoderm saraf tanpa kompetensi saraf.
Dua turunan krista neuralis, mesenkim dan sel pigmen masih dapat ditimbulkan di ektoderm tahap kuncup ekor. Ektoderm yang lebih tua sama sekali tidak memiliki kompetensi untuk melakukan apa pun selain melanjutkan menuju takdir epidermal intrinsiknya (Gbr. 2).
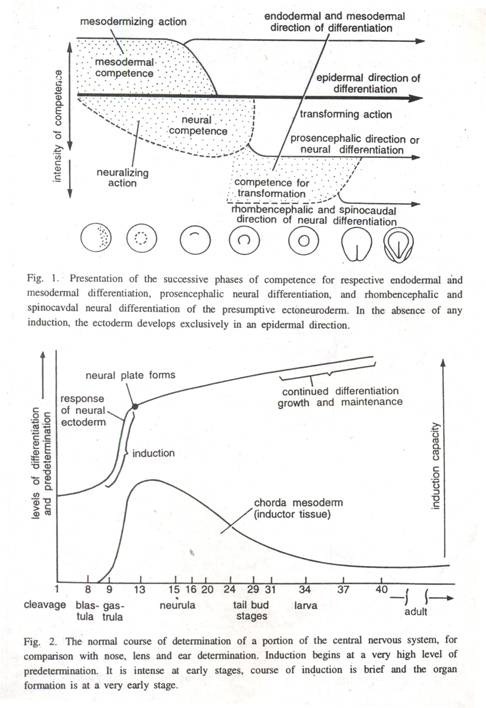
Selanjutnya, neurula epidermis akhir yang tidak lagi dapat diubah menjadi jaringan saraf menjadi kompeten untuk merespons induktor lain di bawah pengaruh masing-masing vesikel mata, otak, dan otak depan. Ini berdiferensiasi menjadi lensa, vesikel telinga dan lubang hidung selama tahap perkembangan pasca-neurula.
Kompetensi biologi molekuler:
Prasyarat untuk kompetensi adalah adanya genom fungsional. Diketahui juga bahwa hingga tahap gastrulasi amfibi, tidak ada transkripsi rRNA yang terjadi. Transkripsi tRNA direpotkan untuk dimulai pada pembelahan akhir (Brown 1964) dan molekul mRNA ditranskripsi sangat sedikit sebelum gastrulasi.
Oleh karena itu, kompetensi terkait dengan timbulnya aktivitas transkripsi genom selama gastrulasi dan produksi protein baru selama gastrulasi juga menciptakan prasyarat untuk kompetensi. Replikasi DNA inti adalah aktivitas yang secara bertahap dilakukan oleh sel-sel embrionik.
Nukleus embrionik tidak menjalankan fungsi normalnya sebelum pola mitosis ini berakhir (Dettlaff 1964). Oleh karena itu, transkripsi dari mitosis sinkron ke asinkron yang terlibat dalam embriogenesis awal membentuk prasyarat untuk kompetensi.
Fenomena induksi dan kompetensi embrio menetapkan fakta bahwa induksi memainkan peran penting dalam perkembangan embrionik. Induksi saraf adalah proses pemicu yang mengatur ektoderm yang kompeten di sepanjang jalur diferensiasi saraf. Peristiwa dan bentuk bahan induktif menentukan luas dan bentuk ektoderm yang terpengaruh.
Area ektoderm yang dineuralisasi seperti itu mengalami pola yang ditentukan sendiri, suatu perkembangan yang khas dari apa yang disebut bidang embrionik. Setiap spesies memiliki repertoar spesifiknya sendiri tentang kemungkinan Cytodiferensiasi. Baik bidang maupun induktor tidak spesifik spesies.
Diferensiasi:
Diferensiasi adalah urutan penuh perubahan yang terlibat dalam diversifikasi progresif struktur dan fungsi sel yang merupakan ciri khas perkembangan. Dari ovum yang telah dibuahi, semua sel muncul melalui pembelahan berulang dan mewarisi anugerah biokimia yang sama, tugas morfogenetik dan fungsi yang sama.
Biasanya proses diferensiasi tidak dapat diubah. Hampir setiap sel embrio yang sedang berkembang berkembang biak dan menjadi berbeda dari kondisi semula maupun satu sama lain. Sel-sel struktur antar sel sebagai serat dalam jaringan ikat, tulang dan matriks tulang rawan, kutikula, dll. membelah untuk menghasilkan sel-sel berdiferensiasi yang terlihat menunjukkan ekspresi morfologis dari fungsi-fungsi khusus sel-sel yang berdiferensiasi. Dalam diferensiasi histologis atau diferensiasi sito, sel mencapai karakteristik biokimia dan fungsional.
Diferensiasi Fungsional:
Baik sel yang tidak berdiferensiasi maupun yang berdiferensiasi mampu melakukan proses metabolisme, gerakan amoeboid, dan reaksi rangsangan refleks lainnya. Selain fungsi-fungsi ini, setiap sel memiliki fungsi khusus, kemampuan morfologis dan biokimia, yang merupakan fungsi umum yang disuperposisikan. Proses ini disebut diferensiasi perilaku. Misalnya, sel-sel hati mengeluarkan empedu.
Diferensiasi kemo:
Diferensiasi kemo berfungsi sebagai pemicu diferensiasi morfofisiologis dan spesialisasi sel di bawah kendali gen. Enzim spesifik atau kombinasi spesifik enzim adalah bagian penting dari mekanisme fungsional sel.
Dengan demikian, dapat dikatakan bahwa diferensiasi adalah produksi pola enzim/protein yang unik. Langkah awal dalam diferensiasi sel dilakukan oleh sitoplasma tetapi tahap akhir tidak dapat dicapai tanpa sintesis protein enzimatik dan struktural, dan berlangsung di bawah bimbingan DNA, RNA, hormon, dan vitamin.
Kontrol Diferensiasi Genik atau Molekuler:
Protein spesifik terlibat, dan diuraikan oleh sel individu untuk mengkarakterisasi pola diversifikasi tertentu. Fenomena ini mungkin disebabkan oleh fakta bahwa genom sel berbeda karena mekanisme replikasi diferensial atau transkripsi DNA, jika identik.
Pengurangan kromatin:
Boveri (1899) pertama kali mengamati penurunan kromatin pada nematoda dan menemukan bahwa dari sel yang paling dekat dengan kutub hewan, bagian dari bahan kromosom tumpah ke dalam sitoplasma selama pembelahan kedua. Pada tahap bersel 32, hanya dua sel yang memiliki komplemen gen lengkap (sel germinal primordial), sedangkan sisanya telah mengalami pengecilan kromatin (dugaan sel somatik).
Transkripsi diferensial DNA:
Dalam operasi transkripsi selektif DNA baik genom total awalnya aktif dan gen individu kemudian ditekan secara selektif atau genom total awalnya tidak aktif dan gen individu kemudian diaktifkan secara selektif untuk transkripsi.
Regulasi aksi gen diferensial untuk diferensiasi:
Pada eukariota, fungsi gen dapat diatur di mana saja antara replikasi gen dan pelepasan rantai polipeptida lengkap dari ribosa sebagai:
DNA→ Transkripsi, RNA→ Terjemahan, Protein
Oleh karena itu, aktivitas gen dapat diatur pada level berikut:
Kontrol diferensiasi pada tingkat genom:
DNA dapat dimodifikasi dengan reaksi alkilasi atau metilasi yang membutuhkan enzim yang ada di dalam sel. Metilasi basis nukleotida DNA apa pun harus dilakukan secara baru di setiap kromosom, setelah replikasi selesai. Karena penurunan atau peningkatan jumlah molekul DNA, kontrol diferensiasi terjadi, di mana jumlah ekstra DNA entah bagaimana mengatur sintesis protein untuk diferensiasi.
Di Xenopus setiap sel normal berisi urutan gen berulang, sekitar 900 salinan gen untuk rRNA, yang disebut gen reduktor. Selama oogenesis, terjadi amplifikasi rDNA sehingga ukuran atau jumlah nukleolus sangat meningkat dan terbentuk sejumlah besar ribosom. Kadang-kadang, penipisan informasi genetik karena penghapusan gen dari salah satu kromosom menyebabkan gen hewan bermutasi.
Kontrol diferensiasi pada tingkat transkripsi:
1. Regulasi gen oleh histon:
Molekul beruntai ganda DNA memiliki gugus asam fosfat bebas di permukaan luarnya dan ini dapat membentuk ikatan yang kuat dengan gugus NH +2 dari asam amino dasar rantai histon. Hubungan erat DNA dan histon ini mencegah DNA dari interaksi dengan zat lain dalam sitoplasma sehingga berfungsi sebagai cetakan untuk produksi RNA. Histon menghambat sintesis RNA prima DNA untuk mengurangi aktivitas DNA polimerase. Dengan demikian, histon berfungsi sebagai represor.
2. Regulasi gen oleh protein asam:
Ini adalah fosfoprotein non-histon, dengan triptofan dan tirosin sebagai konstituen utama. Protein ini tetap terkait erat dengan DNA (kompleks bebas histon) dan dianggap lebih penting untuk histon regulasi gen. Kompleks DNA-histone tetap inert terhadap transkripsi, sehingga protein asam berinteraksi dengan histone dasar, menempatkan histone gen kritis tertentu sebagai promotor sehingga gen dapat ditranskripsi.
3. Regulasi gen dengan heterokromatisasi:
Heterokromatin interfase memiliki beberapa peran spesifik dalam regulasi gen. Misalnya, sintesis protein sangat kurang pada manusia di mana sel darah mengandung banyak heterokromatin terkondensasi, sedangkan pada sel darah putih; sintesis protein sangat kurang karena kurangnya heterokromatin kental.
Kontrol diferensiasi pada tingkat terjemahan:
Selama translasi, pesan yang dibawa oleh mRNA harus didekodekan dan asam amino yang dibutuhkan diambil untuk membentuk berbagai protein, hormon, enzim, dll. menghasilkan diferensiasi dengan memengaruhi jalur sintesis protein. Mekanisme regulasi pasca-transkripsi ada di beberapa langkah yang terlibat dalam pembentukan protein.
Hormon ditemukan lebih efektif untuk menginduksi diferensiasi pada langkah translasi dan mereka dapat memfasilitasi difusi mRNA tertentu keluar dari nukleus, menstabilkan masa hidup mereka di ooplasma, meningkatkan frekuensi mereka mengasosiasikan diri dengan ribosom dan menjadi diterjemahkan dan seterusnya. .
Hormon dapat memengaruhi aktivitas enzim pada tingkat translasi atau menghasilkan efek sekunder atau memengaruhi aktivitas gen kromosom dengan terlokalisasi di nukleus. Selama stimulasi hormonal mitosis diferensiasi mendahului proses diferensiasi terminal.
Menurut Britten dan Davidson (1969) hormon estrogen (Gbr. 3) mempengaruhi dinding rahim dengan meningkatkan protein sel total; pengangkutan asam amino ke dalam sel; aktivitas sintesis protein per satuan jumlah poliribosom; sintesis ribosom baru; ribosom per sel; massa dan jumlah nukleolus; aktivitas dua polimerase RNA; sintesis protein kontraktil; sintesis banyak fosfolipid: sintesis de novo purin dan sintesis mukopolisakarida.
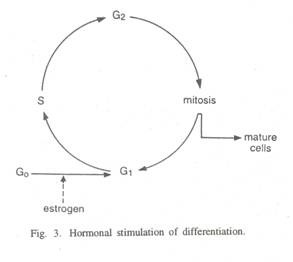
Efek lebih lanjut hormon estrogen pada sel dinding rahim adalah perubahan jumlah protein inti; perubahan rangsangan membran; perubahan metabolisme glukosa, dll.
Tingkat diferensiasi:
Rutter dan Wessells (1967) bekerja pada pengembangan pankreas tikus dan tikus berkaitan dengan sel eksokrin dan endokrin dan mengenali empat tingkat diferensiasi:
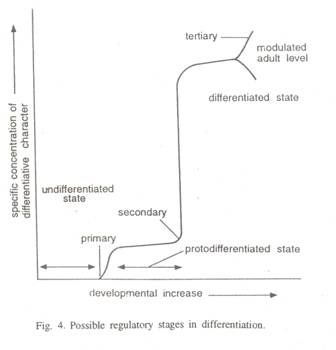
1. Keadaan tidak terdiferensiasi:
Ini terdiri dari konversi sel yang tidak berdiferensiasi menjadi sel dengan karakter pankreas. Pada tahap ini, bahkan sebelum dimulainya histogenesis, protein pankreas spesifik hadir pada tingkat yang dapat dideteksi dan membuktikan bahwa peristiwa ini dapat mengungkap semua gen yang diperlukan untuk diferensiasi pankreas lengkap (Gbr. 4).
2. Status protodiferensiasi:
Peristiwa ini melibatkan konversi dari keadaan protodiferensiasi menjadi keadaan terdiferensiasi. Sel mengalami proliferasi ekstensif, yang berhenti pada sel tersebut, yang mengalami diferensiasi. Sintesis rRNA, perakitan ribosom yang berfungsi atau pengembangan retikulum endoplasma adalah peristiwa utama di negara bagian ini.
3. Keadaan terdiferensiasi:
Ini melibatkan mekanisme yang membuat sel pasti berbeda dari yang lain. Perubahan aktivitas enzim spesifik di antara sel-sel yang berdiferensiasi dapat diamati.
Modulasi keadaan terdiferensiasi mencakup perubahan dalam produksi mRNA spesifik, dan perubahan dalam efisiensi ribosom dalam sintesis protein yang sangat terlambat dalam perkembangannya. Modulasi terjadi sebagai respons terhadap faktor ekstraseluler seperti hormon. Diferensiasi sel induk hampir mirip dengan berbagai tingkat diferensiasi. Mitosis memainkan peran penting untuk pertumbuhan dan juga untuk diferensiasi.
Sel induk:
Sel induk adalah sel yang tidak berdiferensiasi, yang bertindak sebagai bahan sumber untuk jaringan khusus. Sel-sel ini muncul dalam embrio dan tetap ada sepanjang hidup hewan, pada akhirnya bertindak sebagai prekursor untuk jaringan khusus. Sel induk dapat dibedakan dengan pergeseran potensi perkembangan dari kondisi totipoten ke kondisi yang lebih terbatas. Erythropoiesis memberikan contoh yang baik dari sel punca di mana terjadi tingkat diferensiasi yang hampir identik.
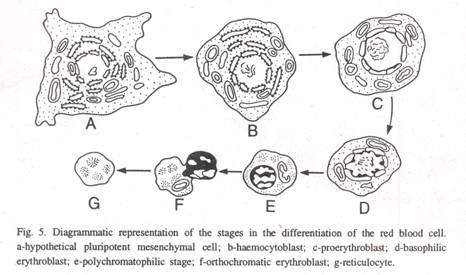
Eritropoiesis:
Ini mencakup serangkaian perubahan bertahap berurutan di mana sel Mesenkim pluripoten menerima rangsangan dan mengarahkan dirinya ke rangkaian eritrositik untuk membentuk sel induk prekursor, yang disebut haemocytoblast. Proses ini disebut sebagai penentuan. Sekarang haemocytoblast menjadi proerythroblast. Proerythrocytoblast ini mengikuti lima langkah yang disebut tahap pematangan dan dapat digambarkan sebagai:
(1) Proerythroblast adalah tahap sintesis RNA aktif dan munculnya protein baru bersama dengan empat pembelahan sel. Kematangan morfologis terjadi dalam enam tahap tertentu di mana perkalian; pertumbuhan dan sintesis hemoglobin berlangsung.
(2) Eritroblas basofilik dicirikan oleh kondensasi kromosom, penurunan aktivitas nuklir, dan produksi hemoglobin.
(3) Tahap polikromatofilik merupakan tahap penting di mana sintesis dan akumulasi hemoglobin meningkat dan tingkat sintesis RNA perlahan menurun.
(4) Eritroblas ortrokromatik adalah sel yang memiliki nukleus tidak aktif dan sitoplasma berisi hemoglobin. Ini tidak mampu pembelahan sel.
(5) Retikulosit matang setelah tiga hari pembelahan sel dan berdiferensiasi menjadi sel darah merah yang berisi hemoglobin. Selama tahap ini, sel mengeluarkan nukleusnya pada mamalia (Gbr. 5).
Interaksi jaringan selama diferensiasi:
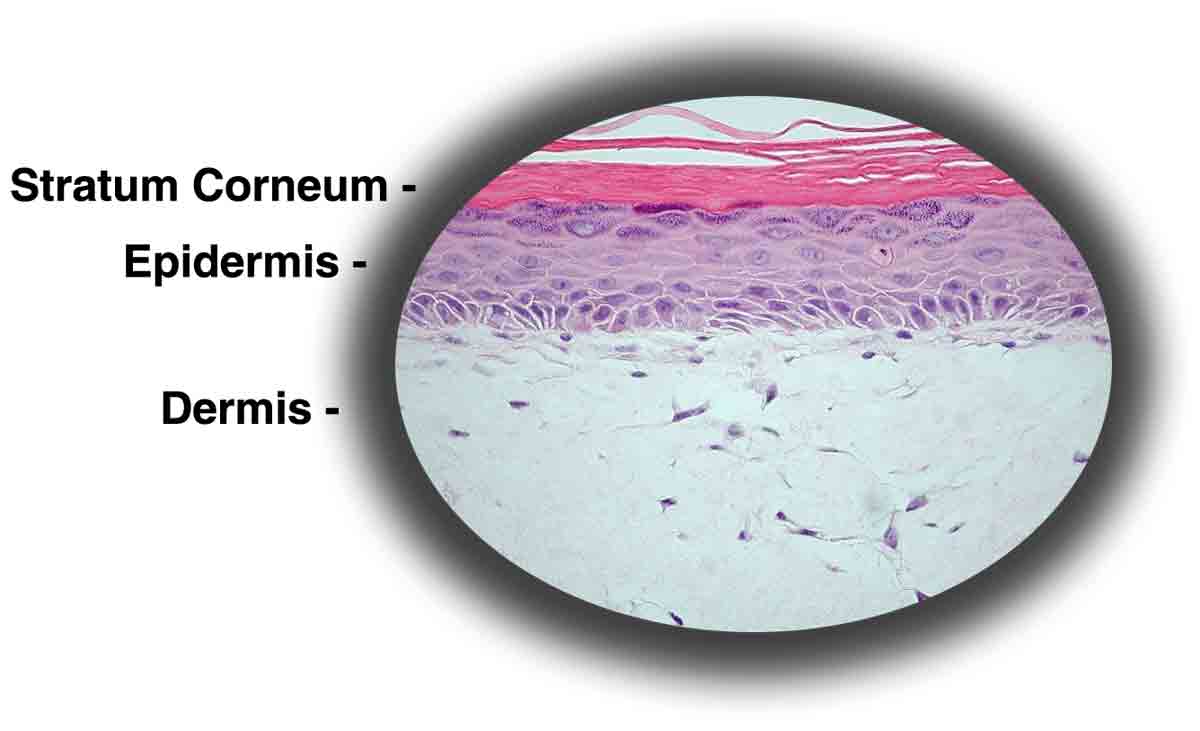
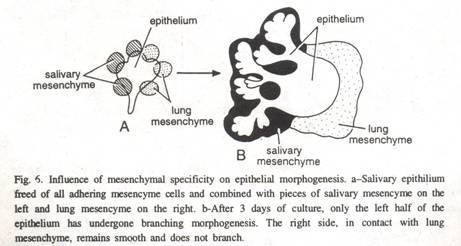
Pada embrio amfibi awal, diferensiasi epitel bergantung pada interaksinya dengan mesoderm. Ketika diisolasi tanpa sel mesenkim, epidermis embrionik kehilangan susunan epitel, yang pada akhirnya menyebabkan degenerasi dan kematian sel.
Dengan sel mesenkim, epitel dipertahankan dan karena diferensiasi ektoderm yang sehat, perkembangan epidermis kulit normal terjadi (Gbr. 6). Perkembangan yang tersinkronisasi dan hubungan intim mesenkim dan epitel menunjukkan sistem kontrol timbal balik yang saling berinteraksi. Kadang-kadang, morfogenesis epitel dimodifikasi oleh mesenkim yang tumbuh, menghasilkan epitel yang mencerminkan asal mesenkim.
Dengan demikian, dapat dinyatakan bahwa sel dan jaringan cenderung ke lokasi akuifer sesuai dengan karakter diferensiasi individu atau kolektifnya atau sel dan jaringan cenderung berdiferensiasi menurut lokasinya dalam sistem yang terorganisir.
Peran lingkungan mikro dalam diferensiasi:
Lapisan sel basal di epidermis terutama terdiri dari sel-sel yang berproliferasi, yang kehilangan keterikatannya dengan membran dasar di bawahnya dan berkerumun ke lapisan superfisial epidermis untuk mensintesis keratin. Sel-sel lapisan basal ini seperti sel punca, yang berdiferensiasi sebagai respons terhadap setiap perubahan kondisi lingkungan. Kedua, sel-sel ini harus terpapar vitamin A untuk memodifikasinya menjadi epitel sekresi mukoid.
Jadi lingkungan mikro sel memainkan peran utama dalam memperbaiki arah diferensiasi yang diambil oleh sel yang tidak terspesialisasi. Jika kita menempatkan dua sel dalam setetes media kultur nutrisi, salah satunya akan terdiferensiasi. Perlahan-lahan, sel lain menerima isyarat dari sel tetangga dan memulai proses diferensiasi (Wilde, 1961).



